|
Genesis: исторические исследования
Правильная ссылка на статью:
Нилогов А.С.
Y-хромосомные предки: к проблеме генетико-генеалогической номинации
// Genesis: исторические исследования.
2024. № 6.
С. 97-166.
DOI: 10.25136/2409-868X.2024.6.43555 EDN: JBURWM URL: https://nbpublish.com/library_read_article.php?id=43555
Y-хромосомные предки: к проблеме генетико-генеалогической номинации
Нилогов Алексей Сергеевич
кандидат философских наук
действительный член (академик), Академия философии хозяйства (Москва); председатель, Южно-Сибирское историко-родословное общество (Абакан); член Совета, Российская генеалогическая федерация; член Региональной общественной организации Красноярского края "Союз краеведов Енисейской Сибири"
655017, Россия, республика Хакасия, г. Абакан, ул. Щетинкина, 23, каб. 23
Nilogov Aleksei Sergeevich
PhD in Philosophy
Current Member (Academician), Moscow Academy of Philosophy of Economy; Chairman of South-Siberian Historical-Genealogical Society (Abakan); Member of Russian Genealogical Federation; member of the Regional Public Organization of the Krasnoyarsk krai "Union of Local Historians of Yenisei Siberia"
655017, Russia, Republic of Khakassia, Abakan, Shchetinkina str., 23, office 23

|
nilogov1981@yandex.ru
|
|
 |
Другие публикации этого автора
|
|
|
DOI: 10.25136/2409-868X.2024.6.43555
EDN: JBURWM
Дата направления статьи в редакцию:
11-07-2023
Дата публикации:
01-07-2024
Аннотация:
В статье рассматривается проблема номинации патрилинейных сопредков, определяемых на основе полногеномного секвенирования мужской половой Y-хромосомы. В результате выявления необратимых мутаций (однонуклеотидных полиморфизмов) становится возможным именование наших далёких пращуров с помощью индексов SNP-маркеров. Ранее обосновав метод «снипования как именования», мы увеличили номинативную ретроспективу по реконструкции филогенетических и генеалогических линий. Благодаря использованию индексальных имён Y-хромосомных мутаций удалось косвенным способом восполнить имена собственные тех сопредков, которые жили ранее сохранившегося архивно-документального фонда генеалогических источников. Речь идёт об ономафикации как процессе воскрешения имён предков, чьи генетические следы возникли в наших Y-хромосомах в виде снип-мутаций, по сути атрибутируя конкретного мужского прародителя, у которого впервые случился необратимый нейтральный ОНП. Молекула ДНК подобно биологическому документу содержит информацию о нашем происхождении на сотни тысяч лет вглубь, являясь генетическим кладбищем удачливых пращуров. Чем больше человеческих Y-хромосом будет секвенировано, тем больше имён собственных мужских предков будет реконструировано на Y-гаплодреве. Несмотря на то обстоятельство, что номенклатура SNPs постоянно меняется, в том числе за счёт синонимизации обозначений в зависимости от разработчиков и лабораторий, непреложным остаётся факт номинативной реконструкции наших филогенетических сопредков посредством масштабного полногеномного секвенирования. На примере генетико-генеалогической (генетеалогической) реконструкции патрилинии автора показаны перспективы комплексного (междисциплинарного) исследования человеческого патрилинейного родства.
Ключевые слова:
Y-хромосома, мутация, генеалогия, ДНК-генеалогия, гаплогруппа, Y-хромосомный Адам, SNP-маркеры, гаплодрево, субклад, филогения
Abstract: The article deals with the problem of the nomination of patrilineal contiguities determined on the basis of genome-wide sequencing of the male sexual Y chromosome. As a result of the detection of irreversible mutations (single nucleotide polymorphisms), it becomes possible to name our distant ancestors using SNP marker indices. Having previously justified the method of "sniping as naming", we have increased the nominative retrospective on the reconstruction of phylogenetic and genealogical lines. Thanks to the use of index names of Y-chromosomal mutations, it was possible to indirectly fill in the proper names of those neighbors who lived earlier in the preserved archival and documentary fund of genealogical sources. We are talking about onomatization as a process of resurrecting the names of ancestors whose genetic traces appeared in our Y chromosomes in the form of snip mutations, in fact attributing a specific male progenitor, who for the first time had an irreversible neutral ONP. The DNA molecule, like a biological document, contains information about our origin hundreds of thousands of years deep, being a genetic cemetery of lucky ancestors. The more human Y chromosomes are sequenced, the more names of proper male ancestors will be reconstructed on the Y-haplodreve. Despite the fact that the nomenclature of SNPs is constantly changing, including due to the synonymization of designations depending on developers and laboratories, the fact of nominative reconstruction of our phylogenetic neighbors through large-scale genome-wide sequencing remains immutable. On the example of the genetic-genealogical (genealogical) reconstruction of the author's patrilineal line, the prospects of a comprehensive (interdisciplinary) study of human patrilineal kinship are shown.
Keywords: Y chromosome, mutation, genealogy, DNA genealogy, haplogroup, Y-chromosomal Adam, SNPs, haplotree, subclade, philogeny
Движение назад во времени в поисках предков может иметь вполне оправданную цель. Эта цель – великий предок всего живого, и мы придём к нему независимо от того, с кого начнём путешествие: со слона или орла, стрижа или сальмонеллы, секвойи или человека. Обратная и прямая хронология хороши каждая для своей цели. Двигаясь в прошлое, мы, независимо от точки старта, придём к моменту единения всего живого.
Р. Докинз [1, с. 19]
Геном белого медведя, пингвина, каймана или гуанако представляет собой экологическое сообщество генов, процветающих в присутствии друг друга. В краткосрочной перспективе ландшафтом совместного процветания выступает клетка. В долгосрочной перспективе – это генофонд. У существ, размножающихся половым путём, генофонд представляет собой ареал обитания генов, которые копируются и подвергаются рекомбинации по мере смены поколений.
Р. Докинз [1, с. 542]
За сотни миллионов лет свершается так много эволюционных преобразований, что предок, выглядящий как рыба, даёт начало потомку, выглядящему как землеройка. А за миллиарды лет – так много, что предок, похожий на бактерию, даёт начало потомку, похожему на меня или на вас.
Р. Докинз [2, с. 204]
ДНК безразличны наши судьбы и имена. ДНК просто есть. А мы пляшем под её дудку.
Р. Докинз [3, с. 173]
Я наугад выбрал 250 мистеров Сайксов из Йоркшира и соседних к нему Ланкашира и Чешира и написал им письма с просьбой прислать материал для исследования ДНК. Посольку и я сам ношу ту же фамилию, мне казалось, что обращение к другим Сайксам не будет воспринято ими как вторжение в личную жизнь. В каждый конверт я вложил щёточку для ДНК и примерно через месяц получил шестьдесят образцов ДНК Сайксов. <…> Однако, когда я окончил расшифровку, результат меня поразил. У доброй половины всех Сайксов из Йоркшира, Ланкашира и Чешира, приславших пробы, структура Y-хромосомы полностью совпадала. Такому удивительному и неожиданному феномену могло быть только одно объяснение: те участники исследования, включая сэра Ричарда и меня самого, у кого структура Y-хромосомы совпала, имели общего предка.
Б. Сайкс [4, с. 19, 21]
Отголоски из прошлого, эхом доносящиеся от наших предков, гораздо легче интерпретировать по митохондриальной ДНК и Y-хромосоме, чем по бесконечно меняющимся хромосомам ядра.
Б. Сайкс [4, с. 173]
…хотя у каждого ныне живущего человека (а также, в данном контексте, у каждого зверя и птицы) на Земле были предки, это вовсе не означает, что у каждого найденного ископаемого существа обязательно имеются современные нам потомки.
Б. Сайкс [5, с. 118]
Несомненно, что ДНК – это физический объект, который в буквальном смысле слова передаётся от поколения к поколению, но в данном случае этот объект важен скорее благодаря тому, что является символом или знаком общности происхождения, о котором он свидетельствует, чем благодаря химическому обмену организма, который он непосредственно контролирует.
Б. Сайкс [5, с. 296]
Архивные записи могут сгореть в пожаре, их могут съесть термиты, уничтожить плесень и сырость, наконец, они могут быть утеряны. ДНК способна заполнить эти пробелы, возникающие в архивах на местах пропавших документов. Она помогает скомпенсировать недолговечность, присущую записям, сделанным с помощью пера и бумаги, но есть множество людей, для которых полное отсутствие каких бы то ни было письменных документов объясняется не стечением обстоятельств, а намеренным преданием каких-то фактов забвению. В этих случаях ДНК не просто полезное дополнение к традиционным методам генеалогии. Она превращается в единственную физическую связь с прошлым.
Б. Сайкс [5, с. 298]
Родство, и только оно одно, должно быть использовано и для реконструкции филогенеза, и для классификации организмов.
В. Хенниг – М. Винарский [6, с. 107]
Мы постепенно приближаемся к воплощению великой мечты – построению универсального древа жизни, связывающего в рамках одной генеалогии все создания, большие и малые, вымершие и ныне живущие.
М. Винарский [6, с. 118]
По мне, о родстве вернее говорит последовательность ДНК. В ДНК накапливаются тысячи мутаций, они возникают независимо друг от друга и не влияют ни на облик, ни на повадки животного. В морфологических чертах, напротив, заключаются средства выживания, поэтому те или иные измерения признаков отражают адаптивные возможности животного. Кроме того, признаки могут быть взаимно увязаны друг с другом – взяв два признака, никогда нельзя быть уверенным в их независимости.
Так как в случае с ДНК мы имеем дело с множественными независимыми и случайно варьирующими признаками, то реконструкции получаются существенно более устойчивыми, чем основанные на морфологических вариациях. И даже больше – на базе ДНК можно получить время расхождения потомков с общим предком, чего никак нельзя сделать по морфологии. Ведь количество изменений в ДНК – это в основном функция времени, по крайней мере если речь идёт о группе родственных видов.
С. Пэабо [7, с. 102–103]
Как можем мы представить события, происходившие миллионы, а во многих случаях и миллиарды лет назад? К сожалению, нельзя расспросить очевидцев – никого из нас не было тогда на свете. Большую часть времени не было не только ни одного говорящего существа, но и ни одного существа, которое имело бы рот и даже голову. Хуже того, животные, которые жили в те времена, умерли и погребены так давно, что от тел лишь немногих из них вообще хоть что-то осталось. Если задуматься о том, что более 99% всех когда-либо живших видов к настоящему времени вымерло, что лишь очень малая их доля сохранилась в ископаемом виде и что ещё меньшую долю от этой доли удается найти, то может показаться, что любые попытки познать наше прошлое изначально обречены на провал.
Н. Шубин [8, с. 11–12]
Все согласятся, что их родословная где-то начинается, но весь вопрос в том, где именно её начало.
Н. Шубин [8, с. 238]
Подумайте, каковы шансы, что, гуляя по какому-нибудь случайно выбранному на нашей планете кладбищу, я обнаружу могилу своего предка? Они крохотны. Что я действительно могу обнаружить, так это то, что все люди, похороненные на любом кладбище – где бы оно ни находилось, в Китае, в Ботсване или в Италии, – в разной степени приходятся мне родственниками. Это можно узнать, исследуя их ДНК с помощью одной из многих продвинутых методик, применяемых сегодня в следственной экспертизе. Я могу убедиться в том, что одни из тех, кто покоится на этом кладбище, состоят со мной в далёком родстве, а другие приходятся мне довольно близкими родственниками. Построенное на основании таких данных родословное древо пролило бы немало света на моё прошлое, на историю моего рода.
Н. Шубин [8, с. 240–241]
Бактерии мыслят группами, штаммами, ветвями генетического древа.
Н. В. Кукушкин [9, с. 105]
Моя любимая точка отсчёта «примечательности» человеческой родословной начинается с эукариогенеза.
Н. В. Кукушкин [9, с. 477]
Древо жизни – это совокупность целых организмов (а не только их геномов). Графически оно в самом деле похоже на дерево, ветви которого, правда, могут иногда и сливаться.
С. А. Ястребов [10, с. 445]
Обычному человеку может быть нелегко совместить с наглядынми представлениями о мире тот факт, что его личный прямой предок – ничуть не менее прямой, чем дед или прадед, – когда-то, пару миллиардов лет назад, был одноклеточным жгутиконосцем, похожим на колледиктиона. Но, судя по всему, это чистая правда. Как сказал герой известной пьесы Григория Горина: «Это гораздо больше, чем факт. Так оно и было на самом деле».
С. А. Ястребов [10, с. 550]
Поэтому на основании принципа естественного отбора, сопровождаемого дивергенцией признаков, представляется вероятным, что от какой-нибудь подобной низкоорганизованной и промежуточной формы могли развиться как животные, так и растения; а если мы допустим это, мы должны допустить, что и все органические существа, когда-либо жившие на Земле, могли произойти от одной первобытной формы.
Ч. Р. Дарвин [11, с. 580]
Новый уровень ДНК-генеалогической (генетико-генеалогической, генетеалогической) верификации поставил под методологический вопрос классическое документально-архивное родословие, так как под родословной понимается прежде всего биологическое (генетическое, филогенетическое) родство.
Написать это исследование меня подвигла книга британского дарвиниста Ричарда Докинза «Рассказ предка», в которой автор рассказал о наших общих сопредках от современности до истока возникновения жизни. Данная работа будет посвящена роду Нилоговых, который реконструирован на основе комплексной генетико-генеалогической методологии.
SNP-мутации (снип-мутации, SNP-маркеры)[1] как способ номинации наших далёких предков – это на сегодняшний день единственная возможность выделить из конечного, но нерасчленённого, числа Y-хромосомных пращуров конкретных биологических носителей этих точечных нуклеотидных замен, которые случились в то или иное время по пути передачи мужской половой хромосомы по прямой линии в цепи «предок – потомок» подобно фамилии, родовому имени[2].
Укоренение филогенетического Y-древа всё ещё нуждается в фундаментальном изучении популяционных генетиков и ДНК-генеалогов (генетических генеалогов, генетеалогов)[3]. В своём исследовании мы будем вести отсчёт от общих предков современных мужчин (Homo sapiens sapiens) с вымершим денисовским человеком (Homo sapiens denisova, Homo sapiens altaiensis). Принадлежа к отряду приматов, семейству гоминид, подсемейству гоминин, роду люди, виду человека разумного, между нашими подвидами существует генетическая связь как по ядерному, так и митохондриальному геномам[4]. В данном случае пойдёт о ядерной ДНК, а конкретно – о геноме Y-хромосомы, референсный образец которой насчитывает более 62 миллионов нуклеотидов (актуальный референс – Т2Т (CP086569.2) [15]). В этой нуклеотидной последовательности – патрилинейная история наших предков, сохранившаяся в виде точковых мутаций, по-другому называемых однонуклеотидными полиморфизмами (SNP). Возникнув случайным образом у наших далёких предков, эти генетические мутации передались нам по наследству в виде биологической родословной, которая подобна родовому кладбищу реплицирует память об удачливых пращурах, кому, выражаясь тафономически, повезло именно таким способом сохраниться в потомках. ДНК содержит в себе огромный массив информации о предках человека, тех персон фамильных линий и их взаимосвязей хромосомного родства, которые так тщательно исследует генеалогия по сохранившимся документам в архивах. В ДНК эти данные сохранились в человеке, они не подвержены уничтожению в результате чрезвычайных ситуаций и не требуют особых условий хранения в отличие от бумажных документов[5].
Если же посмотреть на проблему с точки зрения всего человеческого генома, то окажется, что в нашем хромосомном наборе будет отсутствовать генетический материал многих наших биологических предков. Дело в том, что в процессе мейоза[6] (деления клетки) происходит так называемый кроссинговер (пересечение, перекрёст), то есть обмен участками гомологичных (парных) хромосом, в результате чего количество возможных сочетаний увеличивается в разы, что и обеспечивает такое разнообразие людей. Однако со временем перетасовка в геноме приводит к такому состоянию, при котором теряется предковый генетический материал, размываясь тем сильнее, чем глубже в историю.
Поскольку Y-хромосома лишь частично подвержена кроссинговеру с парной половой X-хромосомой, постольку SNP-мутации в ней гарантируют память о филогенетических предках. С другой стороны, исследователям родословной не стоит обольщаться насчёт того, что их труд не пропадёт даром, который по достоинству оценят благодарные потомки. Законы природы непреодолимы, поэтому нас всех ждёт тотальное вымирание, забвение и распад. Ни генов, ни мемов после нас не останется: ни в популяции/эволюции, ни в культуре/семиосфере мы не сможем наследить на миллионы и миллиарды лет. Однако если человечество станет фактором космической эволюции, преодолевая энтропию[7], то это позволит нам максимально долго продлить следы своего бывания во Вселенной.
Изучение генеалогии – это частная практика борьбы с энтропией забвения в символической сфере памяти. Генеалогическая схема – это упорядоченная структура памяти предков, которая, пронзая прошлое, устремляется в будущее. Наш негэнтропийный (антропийный) долг – воскрешать имена собственные предков (ономафикация) в общем деле воскрешения отцов (патрофикация) и вообще всей жизни (витафикация), а конкретно – посредством секвенирования их геномов в виде цифровых копий. В соответствии с дарвиновской эволюцией[8] живым организмам важно успеть встроиться в биологическую цепь «предок – потомок»; даже немногие homo sapiens sapiens в состоянии выжить в мире культуры, где память не генетическая и энтропийная, а меметическая и негэнтропийная (антропийная).
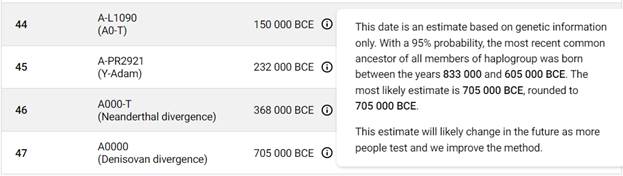
Наша филогенетическая история начнётся с таких основополагающих научныъ понятий, как «most recent common ancestor» (MRCA) и Y-MRCA. Итак, приведём определение первого термина в английском варианте: in biology and genetic genealogy, the most recent common ancestor (MRCA), also known as the last common ancestor (LCA) or concestor, of a set of organisms is the most recent individual from which all the organisms of the set are descended. The term is also used in reference to the ancestry of groups of genes (haplotypes) rather than organisms» [21]. В переводе на русский язык этот термин дословно означает «самый последний общий предок», однако в русскоязычной среде используется эквивалент «ближайший общий предок» (БОП), или «последний общий предок» (ПОП) [22].
Итак, в биологии и генетической генеалогии (ДНК-генеалогии, молекулярной генеалогии, молекулярной истории) самый последний общий предок (MRCA), также последний общий предок (LCA) или сопредок, это из множества ранее существовавших организмов – самая последняя особь, от которой произошло множество современных организмов. Согласно современным эволюционным представлениям вся земная жизнь имеет общее происхождение. Этот фундаментальный принцип играет роль в биологической систематике: согласно кладистическому подходу, любой биологический таксон следует строить таким образом, чтобы он объединял в себе все виды, происходящие от одного общего предка» [22].
Второй важный для нас термин – Y-MRCA: in human genetics, the Y-chromosomal most recent common ancestor (Y-MRCA, informally known as Y-chromosomal Adam) is the patrilineal most recent common ancestor (MRCA) from whom all currently living humans are descended (в генетике человека самый последний общий предок по Y-хромосоме (Y-MRCA, неофициально известный как Y-хромосомный Адам[9]) является самым последним общим предком по отцовской линии (MRCA), от которого произошли все ныне живущие люди)[10] [21].
Итак, Y-хромосомный Адам – это реконструкция ближайшего общего патрилинейного предка для подвида Homo sapiens sapiens, а также для всего вида Homo sapiens, включая неандертальцев (Homo sapiens neanderthalensis) и денисовцев (Homo sapiens denisova), и, естественно, для всего рода homo, включая такие ископаемые виды людей, как гейдельбергский человек (Homo heidelbergensis) и человека прямоходящего (Homo erectus)[11]. Y-MRCA, Y-chromosomal Adam, Y-Adam – синонимичные термины, хотя зачастую под Y-хромосомным Адамом понимают только патрилинейного ближайшего общего предка всех ныне живущих людей. Однако если мы включаем неандертальцев и денисовцев в вид Homo sapiens, то, на наш взгляд, Y-хромосомным Адамом должен считаться патрилинейный ближайший общий предок всех ныне живущих людей и вымерших неандертальцев и денисовцев. Не исключено, что в будущем будут обнаружены другие вымершие подвиды людей или сохранившиеся реликтовые линии, прошедшие бутылочное горлышко, которые также окажут калибровку этого археогенетического понятия[12].
По Brad Larkin [26], имеются данные об общих мутациях вплоть до семейства гоминидов, в частности, с современными шимпанзе, генетически самыми близкими к нам среди отряда приматов. Chimpanzee Last Common Ancestor (CHLCA) – Humans, Denisovans, and Chimpanzee ancestral alleles all match on this last ancestor of humans and chimps. Ларкин приводит конкретный SNP (Hg38) 2784300 на Y-хромосоме, а именно мутацию G > C. CHLCA – это ближайший общий предок современных людей, включая вымерших неандертальцев и денисовцев, а также современных шимпанзе. По аналогии с номенклатурой ISOGG этот сопредок должен образовать гаплогруппу A00000-T, правда, нам ещё не встречалась такая гаплогруппная номинация.
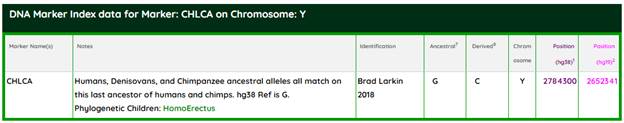
https://www.genetichomeland.com/dna-marker/chromosome-Y/CHLCA [26]

https://www.genetichomeland.com/welcome/dnapedigree.asp?snp=MF203292&Chromosome=Y&snp2=&DB=0 [26]
Нисходящее звено – это ближайший общий предок для современных людей, включая вымерших неандертальцев, и денисовцев, которого Ларкин обозначает Homo Erectus (Human and Denisovan diverge from ancestral allele T found in chimpanzees at this position). Ларкин приводит конкретный SNP (Hg38) 21292569 на Y-хромосоме, а именно мутацию T > G. По номенклатуре ISOGG это должна быть гаплогруппа A0000-T, однако в таком виде термин ещё не вошёл в научный оборот. На сегодняшний день имеется такое рабочее обозначение корня древа (root) для Y-Adam.

https://www.genetichomeland.com/dna-marker/chromosome-Y/HomoErectus [26]

https://www.genetichomeland.com/dna-marker/chromosome-Y/Denisovan [26]

https://docs.google.com/spreadsheets/d/12EwwUDZbwbVx_LswB0PSCS49Zrnb0jnuMS3PWf4gcss/edit#gid=0 [27]
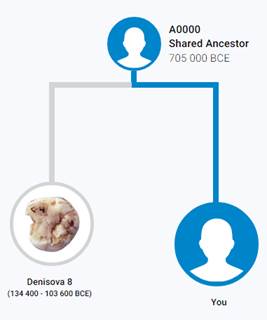
По данным Discover Family Tree DNA дивергенция (разделение) от общего предка денисовцев и современных людей, включая неандертальцев, произошло приблизительно 705 тысяч лет назад (TMRCA)[13] [30].

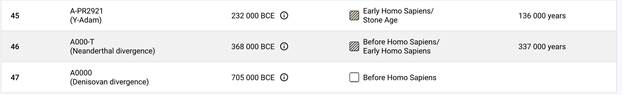
https://discover.familytreedna.com/y-dna/R-Y41478/path [30]
https://discover.familytreedna.com/y-dna/R-Y41478/ancient [30]
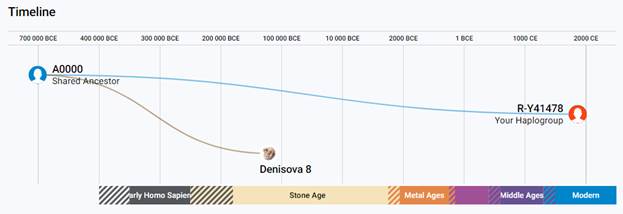
https://discover.familytreedna.com/y-dna/R-Y41478/ancient [30]
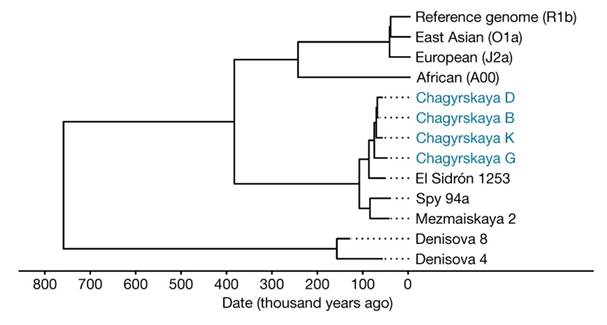
[31, p. 520]
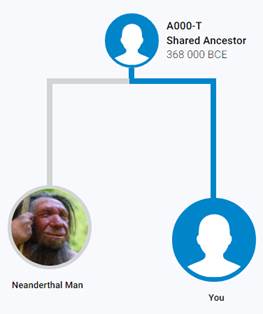
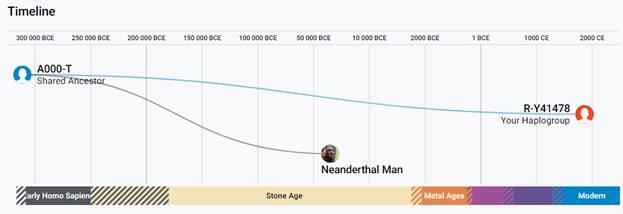
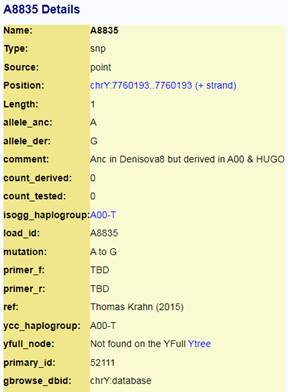
Следующее нижестоящее звено ближайшего общего предка – это Heidelbergensis, от которого образовались две параллельные ветви современных людей и неандертальцев. По номенклатуре ISOGG это гаплогруппа A000-T, которая маркируется SNP A8835: на Y-хромосоме в позиции 7760193 произошла необратимая однонуклеотидная мутация A > G. По данным Family Tree DNA время жизни до ближайшего общего предка (TMRCA) примерно 368000 лет назад в интервале 418000 <–> 324000 лет назад (formed CI 95%)[14].
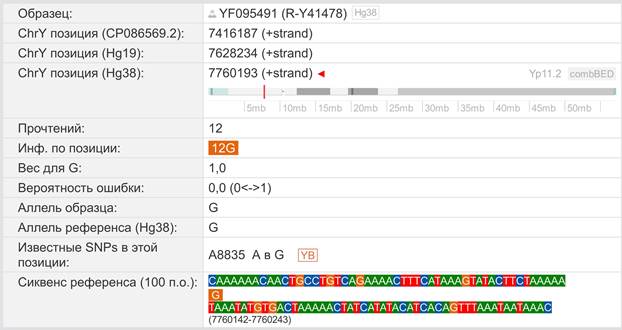
https://www.yfull.com/seq/38/95491/y/7760193/ [32]

https://www.genetichomeland.com/dna-marker/chromosome-Y/Heidelbergensis [26]
https://discover.familytreedna.com/y-dna/R-Y41478/notable [30]
https://discover.familytreedna.com/y-dna/R-Y41478/notable [30]
Методологически поясним этот момент, приведя выдержку из интервью с российским генетиком С. А. Боринской: «Когда при сравнении двух родственных видов в одной и той же позиции генома этих видов стоят разные нуклеотиды, то один из нуклеотидов предковый, другой – эволюционно молодой, заменивший предковый в результате мутации. Чтобы узнать, какой из вариантов предковый, а какой "молодой", используют геном родственного вида (его называют "внегрупповой вид", по-английски outgroup), отделившегося от общего ствола до расхождения тех двух видов, которые изучают[15].
При сравнении генов человека с геномами неандертальца или денисовца в качестве "внегруппового" (outgroup) вида использовали шимпанзе (в ряде случаев гориллу, орангутана, других приматов).
Логика исследования такова: если у человека и денисовца (или неандертальца) в какой-то позиции генома стоят разные нуклеотиды, то тот, который совпадает с шимпанзе, и есть предковый. А тот, который не совпадает – эволюционно молодой, возник уже после отделения линии, несущей этот молодой аллель» [35][16].
Итак, продолжим Y-филогению наших общих предков, начиная по аналогии с Р. Докинзом вести речь о сопредках (сoncestors), всегда подразумевая и женского сопредка, то есть исключая патрилинейную дискриминацию. Мы будем именовать наших сопредков по названиям SNP-маркеров и соответственно гаплогрупп/субкладов[17], которые приняты в Y-филогении. Это научно-номенклатурная SNP-номинация[18] позволит косвенным образом восполнить имена далёких родственников – конкретных носителей мутаций, которые у них однажды возникли[19]. По мере накопления базы данных филогения YTree будет уточняться и расширяться, позволяя именовать как можно больше безымянных пращуров, чьи следы в виде нуклеотидных следов сохранились в Y-хромосоме и подобно биологической родословной реплицируются в потомках. Полногеномное тестирование Y-хромосом мужских представителей нашего рода проводилось в двух американских коммерческих лабораториях – Family Tree DNA (6 доноров) [38] и Nebula Genomics (повторно 2 донора) [39]. Результаты были загружены на международный сайт YFull.com (Y-Chr Sequence Interpretation Service), где они заняли своё уникальное место (соответствующую ветвь) на мировом YTree [40]: А. С. Нилогов (id:YF001593 (Big Y 500), id:YF067279 (Big Y 700), id:YF095491 (NG)); С. М. Нилогов (id:YF068473 (Big Y 700), id:YF089316 (NG)); М. С. Нилогов (id:YF076737 (Big Y 700)); М. М. Нилогов (id:YF112473 (Big Y 700)); Н. А. Нелогов (id:YF079120 (Big Y 700)); А. Н. Нелогов (id:YF110536 (Big Y 700)).
Предок (сопредок) A8835 (A000-T) (+ женщина): у него на Y-хромосоме в позиции 7760193 произошла необратимая однонуклеотидная мутация A > G. По данным FTDNA время жизни до ближайшего общего предка (TMRCA) примерно 368000 лет назад в интервале 418000 <–> 324000 лет назад (formed CI 95%). Филогенетические сопредки узла A8835 – это приблизительно 8 потенциальных биологических мужских предков, которые поименованы следующим образом: A8835, A8836, A8837, A8838, A8845, A8846, A8848, A8852. A8835 (A000-T) – наш общий патрилинейный предок с неандертальцами.

https://www.genetichomeland.com/dna-marker/chromosome-Y/Neanderthal [26]
https://discover.familytreedna.com/y-dna/A-PR2921/ancient [30]
https://ybrowse.org/gb2/gbrowse_details/chrY?ref=chrY;start=7760193;end=7760193;name=A8835;class=Sequence;feature_id=52111;db_id=chrY%3Adatabase [41]
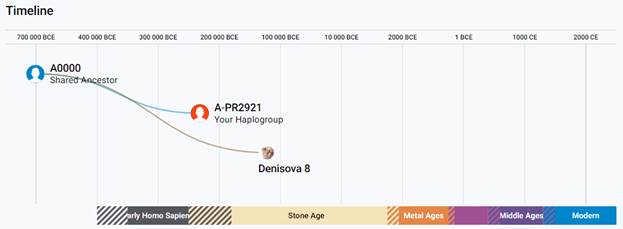
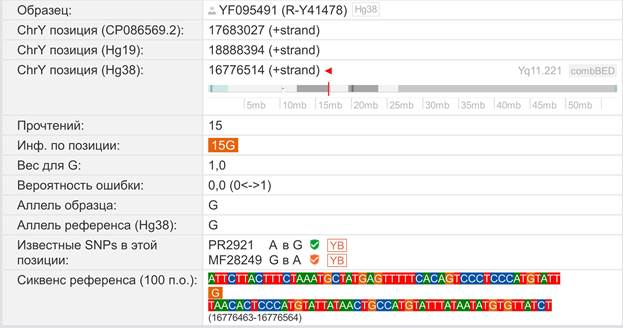
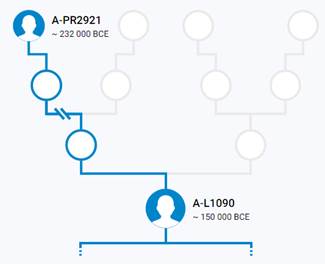
Предок (сопредок) A-PR2921 (A00-T) (+ женщина): у него на Y-хромосоме в позиции 16776514 произошла необратимая однонуклеотидная мутация A > G. По данным FTDNA время жизни до ближайшего общего предка (TMRCA) примерно 232000 лет назад в интервале 268000 <–> 200000 лет назад (formed CI 95%). Филогенетические сопредки узла A-PR2921 – это приблизительно 105 потенциальных биологических мужских предков, которые поименованы следующим образом: PR2921, A8833, A8841, A8843, A8849, A8853, A21321, A21322, A21323, A21324, A21325, A21326, A21327, A21328, A21329, A21330, A21331, A21332, A21333, A21334, A21335, A21336, A21337, A21338, A21339, A21340, A21341, A21342, A21343, A21344, A21345, A21346, A21347, A21348, A21349, A21350, A21351, A21352, A21353, A21354, A21499, A21500, A21501, A21502, A21503, A21504, A21505, A21506, A21507, A21508, A21509, A21510, A21511, A21512, A21513, A21514, A21515, A21516, A21517, A21518, A21519, A21520, A21521, A21522, A21523, A21524, A21525, A21526, A21527, A21528, A21529, A21530, A21531, A21532, A21533, A21534, A21535, A21537, A21538, A21539, A21540, A21541, A21542, A21543, A21544, A21545, A21546, A21547, A21548, A21549, A21550, A21551, A21552, A21553, A21554, A21555, A21556, A21557, A21558, A21559, A21560, A21739, A21740, A21741, A21742. A-PR2921 (A00-T) – наш общий патрилинейный предок после разделения с неандертальцами.
https://www.yfull.com/seq/38/95491/y/16776514/ [32]
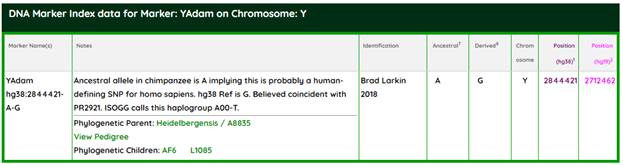
https://www.genetichomeland.com/dna-marker/chromosome-Y/YAdam [26]
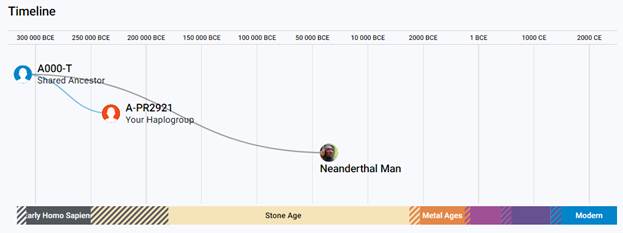
https://discover.familytreedna.com/y-dna/A-PR2921/notable [30]
Важно заметить, что от момента разделения современных людей с неандертальцами по Y-хромосоме, которое случилось приблизительно 368 тысяч лет назад от A8835 (A000-T) до A-PR2921 (A00-T) жили тысячи потенциальных ближайших общих предков для современных людей, которые могут быть филогенетически реконструированы по мере тестирования новых доноров, включая человеческие останки с древней ДНК[20]. Тем не менее, филогенетическая летопись по Y-хромосоме всегда будет неполной вследствие того, что многие линии уже терминировались и продолжают вымирать даже в условиях массового ДНК-тестирования, а часть вымершего генофонда уже не оставила даже костных образцов.
https://discover.familytreedna.com/y-dna/A-PR2921/story [30]
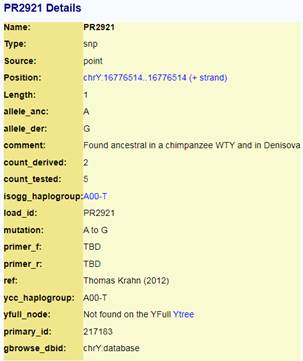
https://ybrowse.org/gb2/gbrowse_details/chrY?ref=chrY;start=16776514;end=16776514;name=PR2921;class=Sequence;feature_id=217183;db_id=chrY%3Adatabase [41]
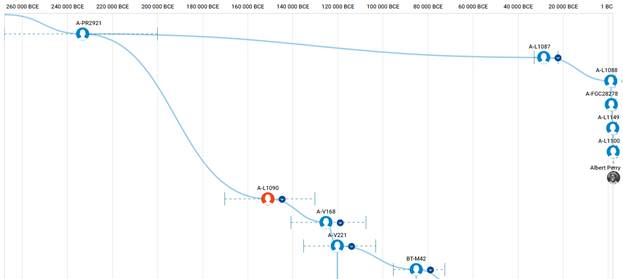
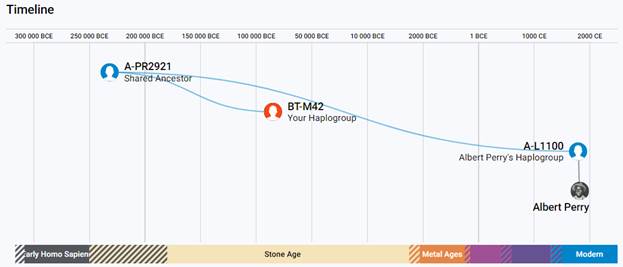
В 2012 году в американской лаборатории Family Tree DNA был протестирован один из правнуков Альберта Перри (Albert Perry), раба из Южной Каролины, который родился около 1820 года. В результате анализа его Y-хромосомы было установлено, что его патрилиния – самая древняя из ранее обнаруженных («the most divergent Y-DNA lineage known today»), корни которой уходят в Камерун. Благодаря этому американцу была выделена гаплогруппа A00. Семья Перри и их дальние родственники из Камеруна происходят от одного предка, который жил около тысячу лет назад. Таким образом, на сегодняшний день они являются нашими самыми дальними родственниками по отцовской линии («they are the most distant paternal line relatives of almost everyone in the world today») [42]. Для правнука Альберта Перри определён субклад A-L1100, который является нисходящим в цепи «предок – потомок»: A-PR2921 > L1087 > L1088 > FGC28278 > L1149 > L1100 > A-FT272432.
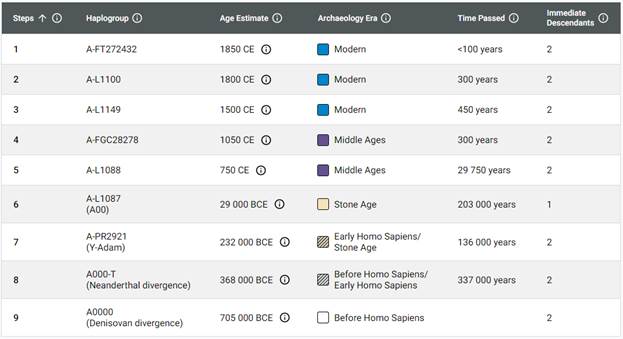
https://discover.familytreedna.com/y-dna/A-FT272432/path [30]
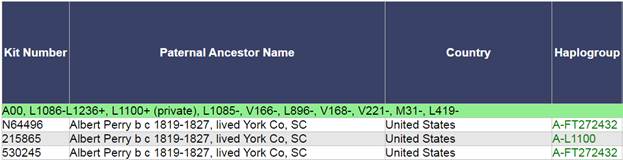
https://www.familytreedna.com/public/Haplogroup_A?iframe=yresults [37]

Albert's son, Clyde Perry, born in 1867, grandfather of the first A00 tester [30]
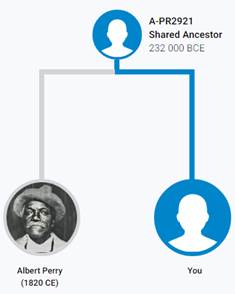
https://discover.familytreedna.com/y-dna/R-Y41478/notable [30]
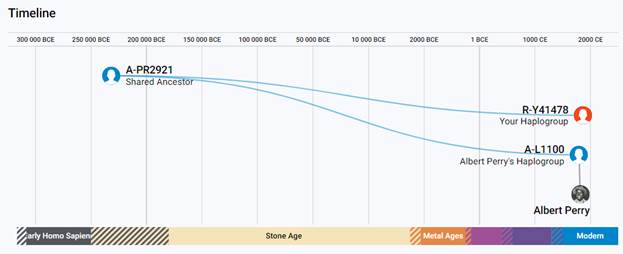
https://discover.familytreedna.com/y-dna/R-Y41478/notable [30]
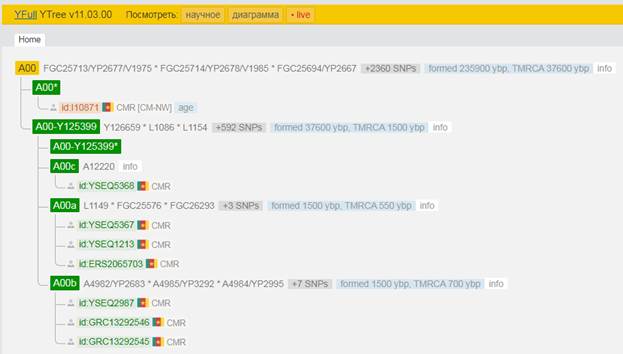
https://www.yfull.com/tree/A00/ [40]
В нашем исследовании мы используем несколько генетических баз данных (Y-DNA database), среди которых наиболее авторитетными являются: YFull YTree [29], Discover Family Tree DNA [30], YSNP YTree от ISOGG [27]. Также имеются китайские варианты – YTree Dnachron [43], YTree Mofang [44], старая версия Y-Haplotree FTDNA [45] и сайт GeneticHomeland.com, на котором размещена утилита Ancestral DNA Marker Pedigree Display [26].
https://www.yfull.com/tree/[21] [40]
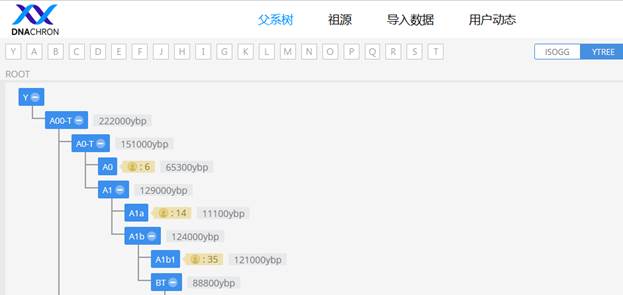
https://www.dnachron.com/ytree [43]
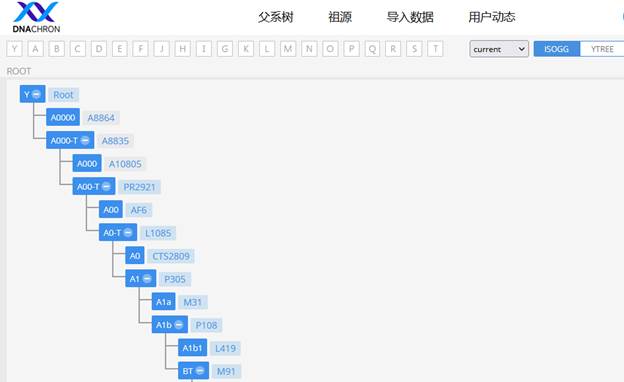
https://www.dnachron.com/isogg [43]
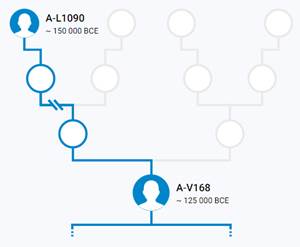
Предок (сопредок) A-L1090 (A0-T) (+ женщина): у него на Y-хромосоме в позиции 3676921 произошла необратимая однонуклеотидная мутация G > C. По данным FTDNA время жизни до ближайшего общего предка (TMRCA) примерно 150000 лет назад в интервале 170905 <–> 131165 лет назад (formed CI 95%). По данным YFull это событие произошло 235900 лет назад в интервале 243700 <–> 228300 лет назад (formed CI 95%), однако YTree у YFull не содержит более глубоких (родительских) снип-мутаций, поэтому по сути гаплогруппу А00-Т (A-PR2921) можно датировать примерно 236 тысячами лет[22]. Поскольку FTDNA и YFull используют разные алгоритмы расчётов хронологии гаплогрупп/субкладов, разные базы с образцами Y-хромосомы и разные SNP-филогении, постольку очевидны различия как в датировании конкретных ветвей, так и в самой структуре Y-гаплодрева[23]. Для нашей Y-филогении мы будем использовать версию YFull YTree[24] [40], так как она включает большее количество ветвлений на гаплогруппы/субклады. Корректировка хронологии для каждого конкретного субклада будет уточняться по мере расширения базы. Ниже приведём путь наших генетических предков от Discover FTDNA[25] [30], а также от YFull в сокращённом виде, далее сосредоточившись на всех сопредках для каждого уровня ветвления YTree.
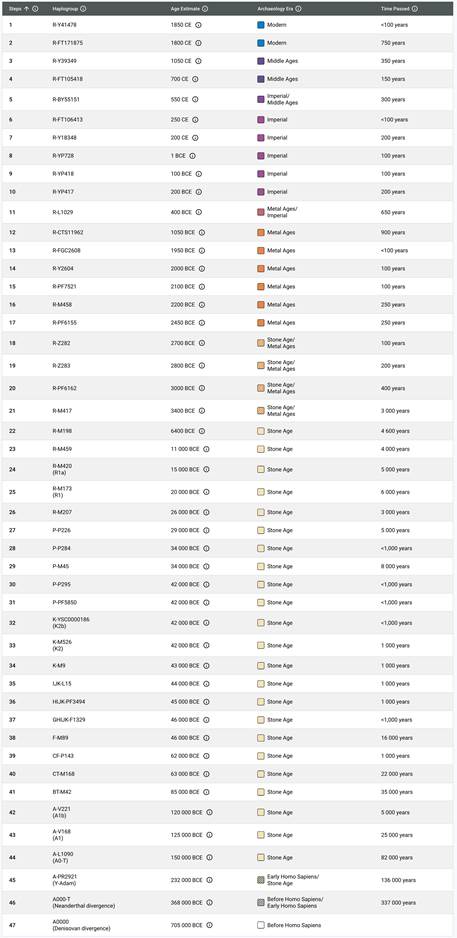
https://discover.familytreedna.com/y-dna/R-Y41478/path [30]
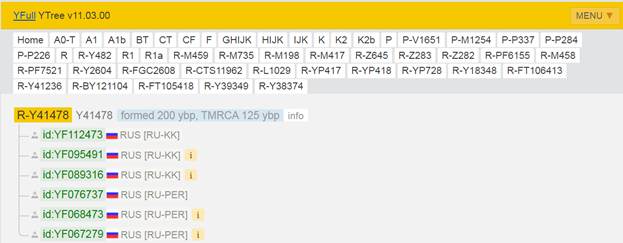
https://www.yfull.com/tree/R-Y41478/ [40]
Филогенетические сопредки узла A-L1090 – это приблизительно 542 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения:FGC26832/YP2013, A2788/YP1957, FGC26530/YP1952, FGC26528/YP1951, FGC26833/YP2014, A2594/YP1741, A2606, A2611/YP1752, A2621/YP1766, A2622/YP1767, A2623/YP1768, A2624/YP1769, A2627/YP1776, A2636/YP1794, A2638/YP1801, A2646/YP1817, A2662/YP1844, A2663/YP1850, A2669/YP1863, A2673/YP1870, A2742/YP1891, A2743/YP1894, A2744/YP1895, A2752/YP1903, A2757/YP1909, A2758/YP1910, A2779/YP1950, A2797/YP2001, A2852/YP2078, A2855/YP2091, A2866/YP2096, A2868/YP2099, A2870/YP2107, A2873/YP2113, A2879/YP2114, A2882/YP2123, A2883/YP2124, A2906/YP2136, A2909/YP2143, A2934/YP2175, A2937/YP2184, A2943/YP2196, A2944/YP2197, A2949/YP2200, A2950/YP2201, A3047/YP1840, A3071/YP1916, A3072/YP1920, A3074/YP1935, A3281/YP2681/V4845, A3341/V5179, A4737/YP1734, A4750/YP1879, A4763/YP2052, A4776/YP2089, AF4, FGC24679/V5187, FGC24700/V2475, FGC24753/YP1711, FGC24755, FGC24761/YP1712, FGC24763, FGC24764/YP1713, FGC24769/YP1714, FGC24772/YP1715, FGC24775/YP1716, FGC24777/YP1717, FGC24781/YP1718, FGC24784/YP1719, FGC24792/YP1720, FGC24794/YP1721, FGC24833/YP1727, FGC25066/YP1731, FGC25299/YP1733, FGC25307/YP1736, FGC25361/YP1742, FGC25366/YP1743, FGC25379/YP1744, FGC25394/YP1745, FGC25401/YP1746, FGC25403/YP1747, FGC25418/YP1749, FGC25420/YP1750, FGC25442/YP1751, FGC25449/YP1753, FGC25450/YP1754, FGC25452/YP1755, FGC25462/YP1758, FGC25464/YP1759, FGC25465/YP1760, FGC25470/YP1761, FGC25472/YP2532, FGC25479/YP1763, FGC25482/YP1764, FGC25488/YP1765, FGC25493/YP1770, FGC25512/YP2555, FGC25515/YP1773, FGC25516/YP1774, FGC25524/YP1777, FGC25529/YP1778, FGC25530/YP1779, FGC25532/YP1780, FGC25533/YP1781, FGC25536/YP1782, FGC25541/YP1783, FGC25543/YP1784, FGC25544/YP1785, FGC25546/YP2573, FGC25550/YP1786, FGC25557/YP1787, FGC25559/YP1788, FGC25563/YP1790, FGC25566/YP1791, FGC25571/YP1792, FGC25574/YP1793, FGC25588/YP1796, FGC25590/YP1797, FGC25596/YP1798, FGC25610/YP1799, FGC25615/YP1800, FGC25623/YP1803, FGC25625/YP1804, FGC25634/YP2621/V1609, FGC25644/YP1805, FGC25649/YP2631/V1662, FGC25656/YP1810, FGC25659/YP1811, FGC25676/YP1815, FGC25695/YP1821, FGC25708/YP1826, FGC25716/YP2680/V1995, FGC25743/YP2699/V2171, FGC25746/YP1833, FGC25748/YP2702/V2201, FGC25754/YP1834, FGC25761/YP2711/V2278, FGC25766/YP2718/V2339, FGC25769/YP2719/V2369, FGC25771/YP1835, FGC25775/YP1836, FGC25781/YP1837, FGC25789/YP1842, FGC25796/YP1843, FGC25803/YP1846, FGC25813/YP1847, FGC25828/YP1851, FGC25829/YP1852, FGC25830/YP1853, FGC25853/YP1855, FGC25860/YP1856, FGC25864/YP1857, FGC25866/YP1859, FGC25872/YP1860, FGC25874/YP1861, FGC25881/YP1862, FGC25887/YP1864, FGC25917/YP2796/V5073, FGC25918/YP1867, FGC25921/YP1868, FGC25926/YP1869, FGC25943/YP1872, FGC25951, FGC26159, FGC26182/YP1875, FGC26187/YP1876, FGC26188/YP1877, FGC26192/YP1878, FGC26197/YP1880, FGC26198/YP1881, FGC26210/YP1882, FGC26223/YP1883, FGC26235/YP1884, FGC26237/YP1885, FGC26238/YP1886, FGC26254/YP1887, FGC26255/YP1888, FGC26257/YP1889, FGC26258/YP1890, FGC26273/YP1893, FGC26276/YP1896, FGC26277/YP1897, FGC26279/YP1898, FGC26280/YP1899, FGC26285/YP1900, FGC26287, FGC26289/YP1901, FGC26298/YP1904, FGC26309/YP1905, FGC26328/V5676, FGC26341/YP1906, FGC26343/YP1907, FGC26356/YP1908, FGC26368/YP1913, FGC26370/YP1914, FGC26374/YP1917, FGC26375/YP1918, FGC26390/YP1921, FGC26405/YP1923, FGC26406/YP1924, FGC26410/YP1925, FGC26423/YP1927, FGC26434/YP1929, FGC26435/YP1930, FGC26439/YP1931, FGC26449/YP1933, FGC26455/YP1934, FGC26462/YP1936, FGC26467, FGC26470/YP1937, FGC26472/YP1938, FGC26476/YP1939, FGC26478/YP1940, FGC26480/YP1941, FGC26484/YP1942, FGC26485/YP1943, FGC26492/YP1945, FGC26493/YP1946, FGC26514/YP1949, FGC26531/YP1953, FGC26539/YP3019/V3152, FGC26541/YP1954, FGC26561/YP1956, FGC26591/YP1961, FGC26593, FGC26606/YP1962, FGC26610/YP1966, FGC26619/YP1968, FGC26620, FGC26627/YP1971, FGC26629/YP1972, FGC26632/YP1973, FGC26706, FGC26737/YP1997, FGC26773/V6456, FGC26779, FGC26799/YP3161, FGC26816/YP3176, FGC26835, FGC26852/YP2021, FGC26891/YP2029, FGC26921/YP3231, FGC26939/YP2039, FGC26959/YP2042, FGC26961/YP2043, FGC26987/YP2050, FGC27006, FGC27007/YP2056, FGC27013/YP2057, FGC27025/YP2061, FGC27037, FGC27110/YP2081, FGC27112/YP2082, FGC27113/YP2083, FGC27115/YP2084, FGC27118/YP2086, FGC27136/YP2087, FGC27141/YP2088, FGC27145/YP2090, FGC27146/YP2092, FGC27153/YP2093, FGC27155/YP2094, FGC27182/YP2095, FGC27185/YP2097, FGC27186/YP2098, FGC27198/YP2100, FGC27200/YP2101, FGC27201/YP2102, FGC27203/YP2103, FGC27206/YP2104, FGC27214/YP2105, FGC27215/YP2106, FGC27217/YP2108, FGC27218/YP2109, FGC27233/YP2110, FGC27234/YP2111, FGC27236/YP2112, FGC27244/YP2115, FGC27247, FGC27249/YP2116, FGC27250/YP2117, FGC27254/YP2118, FGC27255/YP2119, FGC27257/YP2120, FGC27261/YP2121, FGC27262/YP2122, FGC27272/YP2125, FGC27281/YP2126, FGC27293/YP2128, FGC27299/YP3447, FGC27324/YP3457, FGC27337/YP2135, FGC27344/YP2137, FGC27345/YP2138, FGC27348/YP2139, FGC27349/YP2140, FGC27355/YP2141, FGC27358, FGC27360/YP2142, FGC27371/YP2144, FGC27372/YP2145, FGC27375/YP2146, FGC27382/YP2147, FGC27394/YP2148, FGC27408/YP2149, FGC27410/YP2150, FGC27412/YP2151, FGC27431/YP2154, FGC27434/YP3520/V7628, FGC27435/YP3521/V7633, FGC27443/YP2155, FGC27476/YP2161, FGC27477/YP2162, FGC27481/YP2163, FGC27482/YP2164, FGC27486/YP2165, FGC27489/YP2166, FGC27493/YP2167, FGC27496/YP2168, FGC27501/YP2170, FGC27502/YP2171, FGC27506/YP2172, FGC27538/YP2176, FGC27542/YP2177, FGC27555/YP2179, FGC27576/YP2183, FGC27578/YP2185, FGC27579/YP2186, FGC27593/YP2188, FGC27688/YP3605, FGC27698/YP2195, FGC27704/YP2198, FGC27706/YP2199, FGC27710/YP2202, FGC27721/YP2204, FGC27725/YP3626, FGC27728/YP2206, FGC27730/YP2207, FGC27733/YP2208, FGC27734/YP2209, FGC27736/YP2210, FGC27738/YP2211, FGC27743/YP2212, FGC27752/YP2213, FGC27764/YP2214, FGC27769/YP2215, FGC27771/YP2216, FGC27774/YP2217, FGC27782/YP2218, FGC27785/YP2219, FGC27791/YP2220, FGC27792/YP2221, FGC27793/YP3663, FGC27795/YP2222, FGC27802/YP2223, FGC27813, FGC27818/YP3679, FGC27820/YP2224, FGC27826/YP2225, FGC27834/YP2227, FGC27864, FGC27878/YP2232, FGC27884/YP2233, FGC27886/YP2234, L1085, L1098, L1099, L1101, L1105, L1113, L1114, L1116, L1118, L1120, L1121, L1123, L1124, L1125, L1127, L1128, L1130, L1135, L1137, L1142, L1145, L1150, L1155, L1235, M4507, TY2950/AF11, TY2983/M8129, V2247/FGC25756, V2271/FGC25758, V2828/FGC26411/YP1926, V3872, Y125383/BY184360/FGC26002, Y125401/BY184361, Y125423, Y17293, Y49138, YP1737, YP1775, YP1802, YP1806, YP1807, YP1812, YP1814, YP1818, YP1829, YP1830, YP1831, YP1839, YP1854, YP1866, YP1873, YP1911, YP1915/FGC26373/V2657, YP1928/FGC26431, YP1963, YP2130, YP2131, YP2132, YP2153, YP2158, YP2173/A2932, YP2181, YP2189, YP2190, YP2191, YP2192, YP2193, YP2205, YP2230, YP2235/A2979, YP2236/A2980, YP2237/A2981, YP2238/FGC27910, YP2239/FGC27913, YP2240/FGC27917, YP2241/FGC27918, YP2242/FGC27921, YP2243/FGC27922, YP2244/FGC27924, YP2245/FGC27929, YP2246/FGC27936, YP2247/PR4010, YP2252, YP2253/FGC27971, YP2255/A2989, YP2256, YP2257, YP2258, YP2259/FGC28009, YP2261/FGC28010, YP3754/FGC27944, YP3755/A4452, YP3766/FGC27972, YP3771/FGC27983, YP3773/FGC27984, YP3873/A2724, YP3874/A3068, YP3884, YP3885/FGC26501, YP3887/FGC26154, YP3892/FGC27753, YP3894, YP3895, A2554/YP1729(H), A2969/YP3694, A3042(H), A4702/YP1726(H), FGC24743/V6784, FGC24780, FGC24800/YP1723(H), FGC24809/YP1724(H), FGC24824/YP1725(H), FGC24998/YP1728(H), FGC25061/YP1730(H), FGC25094(H), FGC25224(H), FGC25328/YP1738(H), FGC25345/YP1739(H), FGC25505/YP1772(H), FGC25635/YP2622/V1610, FGC25651/YP2634, FGC25742, FGC25782/YP2729/V2456, FGC25902/YP1865(H), FGC26149/YP1874(H), FGC26297/YP1902(H), FGC26409/YP2950, FGC26502/YP1947, FGC26812, FGC26895/YP3222, FGC27714/YP2203, FGC27823/YP3683, FGC27857, L1090(H), L1093(H), TY2945/FGC26646/Y125418, V1957/FGC25709, V2040/FGC25719, Y17301(H), Y17329/V8014, YP1732(H), YP1735(H), YP1838, YP2080, YP3881/FGC26160(H), YP3889/FGC25163(H), YP3890/FGC24818(H), YP3893(H), FGC24722/C106803, A2921/YP2156, FGC25865/YP1858, FGC25944/Y125388, FGC26364/YP1912, FGC26639/YP1978, FGC26656, FGC26738, FGC27543/YP2178, FGC27583/YP2187, L1089, Y14727, Y17292, YP1740, YP1841, YP1996, YP2249, YP2250, YP2260, YP3876/FGC27582, YP3886/A4762, YP3888/FGC27381, YP2228/FGC27841, YP3459/V7462, A2953/YP3638, ALK431/FGC4291.2/V7027, S6863, YP3792/FGC28017, FGC27695/YP2194, FGC27321/YP3455/V7429.
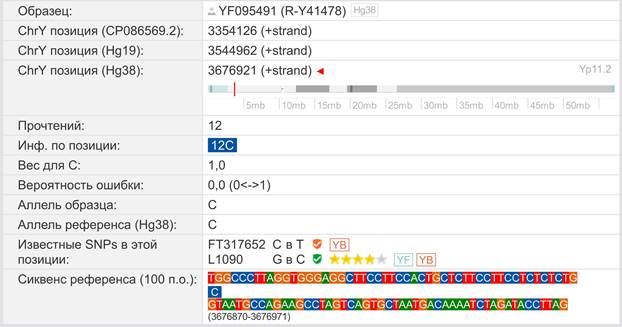
https://www.yfull.com/seq/38/95491/y/3676921/ [32]
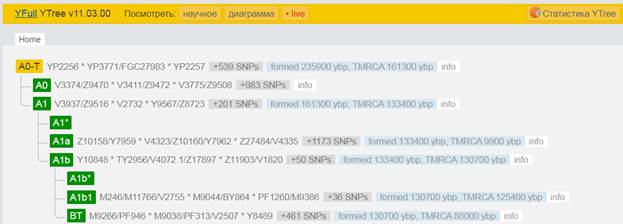
https://www.yfull.com/tree/A0-T/ [40]
https://discover.familytreedna.com/y-dna/A-L1090/story [30]
https://discover.familytreedna.com/y-dna/A-L1090/tree [30]
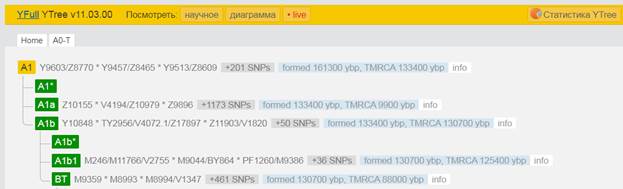
Предок (сопредок) A-V168 (A1) (+ женщина): у него на Y-хромосоме в позиции 15835792 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 161300 лет назад в интервале 169900 <–> 152900 лет назад (formed CI 95%). Филогенетические сопредки узла A-V168 – это приблизительно 204 потенциальных биологических мужских предка, которые поименованы следующим образом:Y9591/Z8755, Y9530/Z8656, Y9786/Z9164, Y9805/Z9192, Y9513/Z8609, Z9719/Y10115, V3682, V2070, Y9977/Z9457, V3832, L986, Y10036/Z9569, YP3853/Z9326, Z9045, Y9747/Z9101, Y9685/Z8912, Z8818/V1522, Y9913/Z9364, Y9870/Z9301, Y9872/Z9304, Y9877/Z9314, Y9697/Z8928, Y10073/Z9613, YP3859, Y9501/Z8584, YP3863, Y9843/Z9265, Y9855/Z9280, Y9814/Z9213, Y9671/Z8883, Z9140/V2613, Y9895/Z9340, Y10169/Z9796, Y9874/Z9310, L1053, Y9718/Z9047, TY2650/Z8834/C126406, Y9972/Z9451, Y9486/Z8553, Y9620/Z8802, Y10075/Z9616, Y9659/Z8866/V2162, V3937/Z9516, Y9998/Z9498, Y9947/Z9415, YP3841, Y9988/Z9474/V3448, Y9863/Z9292, Y1459, Y9643/Z8839, Y9619/Z8801, Y9622/Z8807, YP3852, Y10085/Z9629, Y9807/Z9202, YP3845, Y10220/Z9874, Y9592/Z8757, YP3864, Y9856/Z9282, Z7768, Y10177/Z9808, Z8772, Y9603/Z8770, Y10119/Z9724, Y9800/Z9186, YP3848, Y10195/Z9830, A4698/YP3839, Y9614/Z8792, V174, L985, TY2932/M9072/FT227772, A4709/Y13318, V161/V161.1/V161.2, Y9956/Z9426, Y9558/Z8712, Y9808/Z9203, Y9898/Z9344, Y10061/Z9601, V3624, V2980, Y9457/Z8465, YP3860, Y9910/Z9361, Y9567/Z8723, Y9873/Z9309, L1009, Y9569/Z8725, Z8529, Y10219/Z9873, Y9909/Z9360, Y9950/Z9418, Y10188/Z9820, Y9969/Z9446, Z9656, YP3857, Y13319, Z8830, Y14726, Y9583/Z8745, Y10201/Z9840, Y9669/Z8880/V2359, Y9759/Z9119, Y9661/Z8868/V2174, YP3849, Y13320, A4826/YP3865, YP3858, L1004, Z9582, Y9974/Z9454, V250, Y10090/Z9636, Z9661/Y10106, V171, L1084, Y10019/Z9545, V3711, L1005, Y9623/Z8808, YP3866, YP3891, Z9116, YP3867, Y9904/Z9352, Y9986/Z9467, Y9683/Z8907, YP3842, Y9846/Z9268, V2349, YP3862, V3222, Y9953/Z9420, V1793, Y9713/Z9033, Y9886/Z9327, Z8845/V1808, YP3844, YP3850, YP3846, A4825/YP3861, Y9624/Z8809, V241, Y10068/Z9608, V2842, YP3856, L1002, Y9647/Z8844, Y9680/Z8903, Y9785/Z9163, Y9635/Z8824, Y9793/Z9178, Y9712/Z9032, Y9735/Z9085, Y9971/Z9450, Y10040/Z9574, A4739/YP3847, Y9824/Z9235, Y10039/Z9573, Y10197/Z9832, Y9854/Z9279/C134528, Y9756/Z9114, L989, YP3840, Y9692/Z8923, V2732, Y9639, Y10168/Z9795, V4189, V3653, Y9799/Z9185, A4719, Y10053/Z9590, V2843/Z9173, Y9905/Z9353, YP3868, Y10156/Z9774, Y10034/Z9567, V3315, YP3851, Y9490/Z8559, V2368, Y9798/Z9184, Z9709, V168, A4733, P305, Y9794/Z9179, Z8743, Y9543/Z8686, L1112, Y9456/Z8464, V1130, Y9694/Z8925, V238, Y9768/Z9128, YP3855, YP3854, V1360, YP3843, V3955, V167, Z9072.
https://discover.familytreedna.com/y-dna/A-V168/story [30]
https://www.yfull.com/tree/A1/ [40]
Предок (сопредок) A-V221 (A1b) (+ женщина): у него на Y-хромосоме в позиции 7721262 произошла необратимая однонуклеотидная мутация G > T. По данным YFull это событие произошло примерно 133400 лет назад в интервале 139000 <–> 127800 лет назад (formed CI 95%). Филогенетические сопредки узла A-V221 – это приблизительно 53 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: Y8300, Y8278, Y8283, Y10840, Y10848, Y8298, Y10883, Z11903/V1820, Y8302, Y8288, Y10865, Y10854, Y10856, Y8297, Y8294, Y8293/Z17899, Y8281, TY2957/V4076/Z11917, Y10860, Y10869, Y8289, Y8286, Y8290, V221, Y10882, Y8301, Y10844, Y8303, Y10877, TY2952/V3785/Z11893, Y8894, TY2956/V4072.1/Z17897, Y8299, Y8287, Y8292, Y8280/FGC24622, V2590, P108, Y10870, Y8284, Y8282, Y8296, L1013, Y8285, Y8291, Y8279, Y10857, Y10864, Y10863, Y9420, Y8295, Y10850, Z11900/Y8277.
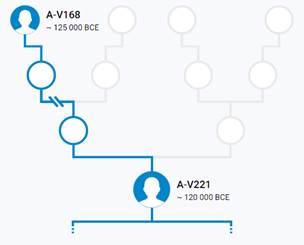
https://discover.familytreedna.com/y-dna/A-V221/story [30]
Предок (сопредок) BT-M42 (+ женщина): у него на Y-хромосоме в позиции 19704954 произошла необратимая однонуклеотидная мутация A > T. По данным YFull это событие произошло примерно 130700 лет назад в интервале 136400 <–> 125100 лет назад (formed CI 95%). Филогенетические сопредки узла BT-M42 – это приблизительно 464 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения:Y10839, M9099/PF674, Y9417/Z17337, P97, L1060/PF1021, M9123, M9298/V4213/Z12093, M9373, M9409/CTS12197/PF1314, Y6870/Z17386, M9196, M9189, M9362, M9411/PF1315, Y10884, M9338/PF1064, M9238, M9130/PF708, L1061/PF1101, M9139, PAGE65.1/SRY1532.1/SRY10831.1/PF6234/SRY10831, M9216, M9244, M9284, V187/PF1403, Y10881/Z17352, M9115/PF687, Y8327/Z12129, M91, Y10852/Z17356, Y30501/Y22371, TY2987/M9404, M9003, Z17385/Y8324, M42, M9365/PF1218, PF280/M11755/Z40388, M9410, M9270/PF952, Y17291, Y8320, M9086/PF648, M9397, M9285, M9331/PF1057, L978/PF93/Z17343, Y10843, Y8308, M9057, M9301/PF1015, Y9452/Z17342, M9001, M8977, Y10861, M9408/PF1296, M9041/PF319, Y1547_2/A4807_2/A4807, Y10845/Z17354, M9179, M9204, M9292/PF995, M8954/V1158, PF1042/Z40407, M8994/V1347, M9195, L962, M9103/PF679, Y1577, L969, L418, Y10849/Z17349, M9286, L1062/PF302/V2352, Y9419, PF632/Z40404, M9295/PF1000/V4130, M9075, M9254, M9231/PF876, PF1406/V102, Y10841, Y8489, M9335/PF1060, M9317, M9094/PF671, M9199/PF834, PF793, M9366, M9165, M299, M9136/PF724, M9353, Y8330/Z17390, M9152/V3226, M8949, M9166/PF785, M9239, M9282/V3904, M9380/PF1256, M9140/V3002, M9121, M9038/PF313/V2507, M8955/PF12, M9319, M9291/V4025, Z17366/Y8309, Y1546_1/A4808_1, M9389, M9017/PF282/V1730, Y10851, PF28, M9203/PF837, M9269/V3636, Y10867, M9104/PF680, M9102, M9421/A5289, L440, M9009/V1561, M9312, M9198, M8999, Y10871/Z17357, M9352/PF1100, M9303, Y9394/Z17344, M8952, M9357/PF1209, M9346, Y10885/Z17362, Y8322, M9289/PF988/V4007, Y10846, M9005/V1506, Y8306, M9347, V29/PF1408, M8973/PF211, M8972, M8971, PF175, M9112, M9368, M8960/PF200, M9021/PF288, M9261/PF931/V3347, M9257/V3304, M9334, PF809/M11773/Z40393, M9224, M9214, M9148/PF744/V3107, L1071/M8945, Y8325, Y9422, Y7547/Z17388, M9054, M9210, M9343/PF1084, Y10876, M9325/PF1052, M9110/PF684/V2561, M9157/PF766, M9300, PF1027/M11779/Z40394, M9255/PF925/V3297, M9128, M9236, M9143/PF732/V3037, Y8314, M9174/S1572, M9133/PF715, M9249, PF1405/V216/M8953, M9126/PF703/V2821, M9097/PF672, M9077, Y9449, V202/PF1404, M9370, M9221, M9374/Z4690, Y10873, L604/PF1243, Y10875, Y8317, M9117/V2634, M9263, M9252, M9138/Z12034, M9176, M9192, Y9451, M9322/PF1049, M9227, M9010, M11781/Z40396, M8947/V1015, M9137/V2952, M9011, M9004/PF270, PF1201/Z40408, M9271, M8980/PF229, M8997/PF260/V1395.1, M9069/PF635, V2315/M11756/PF301/Z40389, Y1546_2/A4808_2/A4808, M9398/PF1279, M9235/PF886, M9114/V2579, Y10858, M8967/Z11946, M9200/PF835, M9188, M9100, M9356, Y10866, M9399/PF1283, M9246, M9305/PF1022, M9027, Y9393/Z17339, Y8319, M9095, M9251/PF913, V8013/Y8326, L970/PF1065, Y8323, M9042, TY2964/M9304, Y8488, Y9392/Z17336, M8983/PF230, Y10837, M9045, M9159/PF767, Y26760/A3036, M9131, M9191, M9420/A5288, M9262/PF932/V3357, M9341/PF1072, M9180, M9405, V59/PF1411, M11752/Z40385, M9155/PF762, PF507, M9146, M9220, M9228, M9187, M9344, M9037/PF311/V2465, M9169, M8961/PF201, M9223/PF865, M9315/PF1033, M9232/PF880, Y10855, M9129/PF707, M9068, M9116/PF688, L977, M9348/PF1093, Y9131, M9230/PF870, M9253/PF914, Y10859/Z17365, PAGES00026/M9336/PAGE26, M9242/PF899, M11760, M9218/PF860, Y10872/Z17345, M9019/PF286/V1813, M9287, M8993, M9070, Y11581, Y10838, M9372, M9265/V3546, PAGES00024/M9160, M9349, TY2727/M9043, M9226/PF869, M9425, M9217/PF857, M9156/PF764, M9197, M9036/PF308, M9107, M8970/PF208, PF592/Z40400, M9396, M9080, M9376, M9202, L438, M9173/PF794, M9290/PF989, M9237/PF890, M9293/PF997, M9124/PF701/V2804, M9002/PF267, M9234/PF885, M9318/PF1039, M8986, V31/L413/PF1409, Y8321/Z17373, Y8318, PF1407/V21/M8969, M8985/PF232, Y10862/Z17372, M9006, Y8311, M9025/V2000, M9020/PF287, Y10842/Z17340, M9367, M9142/PF731/V3032, M9074, M9145/PF733/V3063, M9109, M9321/PF1045, M9083, M9127, M9377/PF1241, M9178, Y10888, M9296, M9032/PF304/V2397, PF161, M9177, M9215/PF847, M9056, M9000/V1456, M9081, M9361, Y10874, M9297/PF1003/V4201, Y8305, TY2992/M9412/FT227766, Y8313, M9311/PF1030, M9277/V3795, M9031/V2319, PF699/M251/M9122/V2760, M9182, M9326, M9039, M9046/PF324, M9379/PF1253, K61, M9360, L1220/M9212, PAGES00081/M9118/V2656/PAGE81, M8976/PF215, Y8329/Z17389, Y8316, M8958/PF196, Y8310, M11754/Z40387, V235/PF1410, M9310, L957, M9151, M9390/PF1262, M9354, M8951, M9193, M9280, M9288/PF985/V3998, PF918/Z40405, M9125, M9260/Z12079, Y9418, M9076, M9066, M9258, M9030/V2318, M9026/V2167, M9359, Z17371/Y8487, M9369, M9340, M9278/PF969, TY2954/M9272, M9393, M8956/PF14, M9065/PF351, M9406, M9163/PF777, Y8312, M9323/PF1050, M9327, M9111, M9417, M9328/PF1053, M8957, Y10847/Z17359, PF236/Z40397, Y9450, M9306, Z17334/Y8304, M8979/PF226, M9089/PF653, M9015, M9064/PF350, M94/PF1081, M9028/PF298/V2209, M9240/PF896, Y8315, PF1126, M9245, M9225/PF868, M9034/V2437, Y10879/Z17341, M9141, M9008/V1530, M9266/PF946, M139, M9098, M9400/PF1284, Y8307, Y10878, Y10853, M8959/PF198, M9375, Y15547, M9316/PF1034, Y10880/Z17348, M9172, TY2967/M9329, Y9421/Z17355, V64/PF1412, M9135, M9248, M9302, M9087, M9175, M9213, M9050, M9113, Y10868, M9394/PF1271, PF1196/FT227729, M8968/PF207, M8988, M9382/PF1257, M9105, PF1247/Z40409, M9219/CTS7503, V1219/M11753/PF243/Z40386, M9209, PF917, M9016, PF601, PF1318, Y10889/Z17346, PF1143, M9283/PF973/V3916, M9267/PF948/V3601.
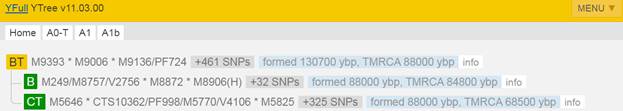
https://www.yfull.com/tree/BT/ [40]
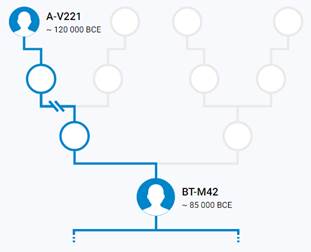
https://discover.familytreedna.com/y-dna/BT-M42/story [30]
https://discover.familytreedna.com/y-dna/BT-M42/notable [30]
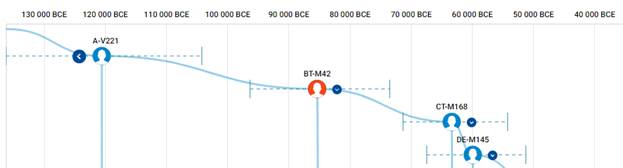
https://discover.familytreedna.com/y-dna/BT-M42/tree [30]
Предок (сопредок) CT-M168 (+ женщина): у него на Y-хромосоме в позиции 12702062 произошла необратимая однонуклеотидная мутация C > T. По данным YFull это событие произошло примерно 88000 лет назад в интервале 92500 <–> 83600 лет назад (formed CI 95%). Филогенетические сопредки узла CT-M168 – это приблизительно 328 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения:M5764/CTS9828/PF964/V3758, M5600, M5753/CTS9458/PF947, PF629, M5757/CTS9555/V3641, M5785, M5617/PF274, M5588/PF210, Y1587, M5636, M5628, Y1544_1, PF15, Y1488, PF192, PF86, M5728/CTS7933, M5631/PF292/V1878, M5691/PF779, Y1455, PF38, M5801, M5754/V3623, Y1538, Y1499, Y1480, Y1450, M5762/CTS9722, M5593, PF6720, M5780, Y1447, M5661/CTS2842, M5808/CTS11358, Y1452, M5783, M5584/CTS543/PF206, M5653/CTS2077/PF657, M5742/PF904, Y1528, M9022/V1863, Y1506, PF636, M5743/CTS8542, Y1591, M5749/V3317, M5632/V2175, M5821/PF1269, Y1819_2/A5213, Y1470, M5788, M5640/PF318, M5804/CTS10946, M5705/CTS6327/PF811, M5768, M5621, M5811/PF1238, M5679/CTS4364, PF1418/V52/M5721, M5648, Y1462, M5720/CTS7482, Y1443, M5652/PF652, Y1567, M5597/CTS1217, M5598/CTS1254, M5771/V4162, M5736/CTS8243/PF891, PF1316, M5669/CTS3431, M5576/CTS125/V1052, M5724/PF866, M5708, PF143, M5818/CTS11991, M294, Y1438, M5620, M5671/CTS3662/PF704/V2824, M5700/CTS6252, M5656, Y1469, M5794/PF1092, PF500, PF328, M5618, Y1473, M5639, PF74, M5689, Y1491, M5690/CTS5318, Y1575_1/A5156_1, M5760/PF954/V3648, M5651/CTS1996, M5763/CTS9760/PF961/V3728.1, Y1454, M5819, M5676/PF720, Y1449, Y1579, M5777, M5729/CTS7936, M5659/PF667, M5776/PF1029, M5687/CTS5019, M5694/CTS5532, M5786/PF1061, M5641, M5781/PF1040, Y1472, M5809/PF1237, Y1585, M5756/PF951, M5751/PF937, M5630, M5590/PF216, M5778/PF1031, M5758/CTS9556/V3642, M5615/PF269/V1494, TY2981/M5803, M5706/PF815, Y1485, M5675/PF719/V2901, M5670/CTS3460, M5649, Z17702/Y7546, Y1507, M5784/PF1059, M5607, M5688, M5583/CTS423, Y1464, Y1440, M5605/V1325, Y1474, M5746/CTS8709, M5790, PF342, Y1489, M5741, Y1471, Y1593, M5802, M5642, PF1239/Z40572, CTS9948, PF1420/V55, Y1527, Y1482, Y1475, CTS10362/PF998/M5770/V4106, Y1476, M5646, PF1414/V9/M5585, M5715/CTS6907/PF833, Y1496, M5727/CTS7922/PF875, M5707/CTS6383, Y1586, M5730/BY14943, Y1525, M5766/V3908/L1480, M5709, Y1451, M5745/CTS8608, M5698/PF796, Y1581, Y1817, Y1492, M5602/PF246, M5797/PF1098, M5739/PF898, M5622, Y1483, M5682, M5732/CTS8089, Y1490, M5633/V2216, Y1493, M5662, PF110, M5609, M5752/CTS9296, M5814, M5713/CTS6800, M5813, M5712, M5717/PF844, PF970/V3858, M5816/CTS11827, Y1465, PF1417/V41/M5695, Y1571, M5832/PF1333, Y1508, M5738, Y1559_1, CTS5248, M5775, M5678/PF725, Y1444, M5718/CTS7257, M5595/CTS1181, M5660, Y1448, Y1580, M5699/PF803, M5711/PF821, M5613, Y1568, Y1552_2, M5826, M9150/PF750, M5619/PF278/V1540, M5647, M5606/PF256, M5664/CTS3120/PF683, M5578, M5812, M5697/CTS5746, Y1509, PF1276, M5716/PF840, M5800/PF1203, M5765, Y1446, M5614/PF266, M5825, M5692, M5767/CTS10110, Y1526, M5587/L1462, Y1467, Y1819_1, M5616/PF272, M5750/CTS9014/V3337, M5737/PF892, PF1415/V226/M5603, PF134, M5805/PF1227, M5612/V1431, M5645, PF228/Z40571, M5772/CTS10512, Y1569/L1492, M5748/CTS8980/PF928/V3310, M5735/CTS8166, M5792/PF1088, M5798, M5626, M5719/PF850, Y1594, PF6718, M5625/V1653, M5791/PF1080, Y1456, M168/PF1416, Y1457, M5726, M5830/CTS12633/PF1329, Y1461, M5796/PF1097, M5624, L1028/CTS4368/M5680, M5684/CTS4740/PF751, CTS7295/PF848, M5782/PF1046, Y1497, PF1016, Y1590, Y1559_2, M5665/CTS3216, M5657, PF1413/V189/M5577, Y1505, TY2730/M5638/PF316, M5831, M5683/CTS4650, Y1599, M5601, M5747, M5722/CTS7517, M5759, Y1791, CTS5262, M5723/PF862, M5589/PF212, M5591/PF223, Y1544_2, M5714/CTS6890, M5725/CTS7741/PF867, Y1495, CTS12325, M5650, M5582/CTS401/PF202, Y1503, Y1460, Y1498, Y1494, M5599/PF234, M5627, M5810/CTS11408, M5681, M5774, PF154, M5629, CTS109/M8948/V1043, PF1205, PF165, M5795, M5817, M5610/V1401, Y1441, CTS5457, Y1458, M5655, M5611/PF263, CTS11575/PF1245, M5822, M5608/PF258, CTS2711, M5686, PF1337, Y1524/FGC24493, PF137, M5769/PF996.
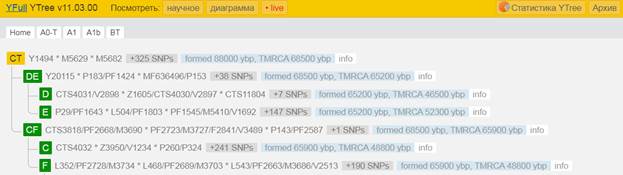
https://www.yfull.com/tree/CT/ [40]
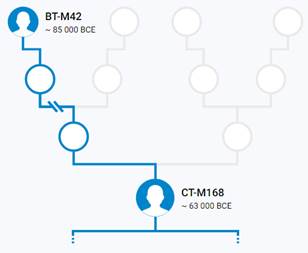
https://discover.familytreedna.com/y-dna/CT-M168/story [30]
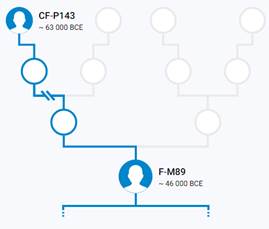
Предок (сопредок) CF-P143 (+ женщина): у него на Y-хромосоме в позиции 12077161 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 68500 лет назад в интервале 71800 <–> 65200 лет назад (formed CI 95%). Филогенетические сопредки узла CF-P143 – это приблизительно 4 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: M3711/CTS6376/PF2697, CTS3818/PF2668/M3690, PF2723/M3727/F2841/V3489, P143/PF2587.
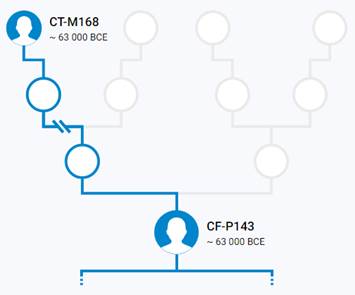
https://discover.familytreedna.com/y-dna/CF-P143/story [30]
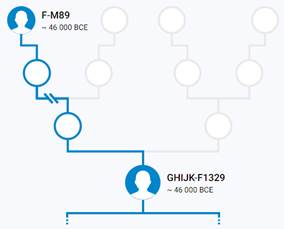
Предок (сопредок) F-M89 (+ женщина): у него на Y-хромосоме в позиции 19755427 произошла необратимая однонуклеотидная мутация С > Т. По данным YFull это событие произошло примерно 65900 лет назад в интервале 69100 <–> 62700 лет назад (formed CI 95%). Филогенетические сопредки узла F-M89 – это приблизительно 193 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: M3699/CTS4737/PF2680, Y1804, PF2732/F2993/M3737/V3940, L882/PF2745/M3749, Y1800, CTS4969/PF2682/M3700, PF2637/M3672, P151/PF2625, PF2739/M3743, Y1811/FGC2054, Y1758/FGC2069, F3584/M3768/PF1916, M3693/CTS4139/PF2672, YSC0001298/PF2620/F1302/M3656, CTS12027/PF2768/M3763, M3772/CTS12662/PF2776, M3730/CTS9372/PF2725, PF2651/F1704/M3675, PF2624/M3659, L352/PF2728/M3734, PF2615/M3652, P159/PF2717, M3692/CTS3996/PF2671, M5685/CTS4838, PF2712, PF2747/M3750, L468/PF2689/M3703, PF2647, PF2589/V186/M3637, P145/PF2617, P138/PF2655, M3666/PF1580, P135/PF2741, P148/PF2734/P148.1/P148.2, CTS11726/PF2765, PF2591/M3639, Y1801, PF2694/F2245/M3709, M3760/CTS11471/PF2764, PF2683/F2048/M3701/V3268, Y1806/FGC2056, TY3619/CTS7981/PF2710, M3725/CTS8985/PF2721, CTS9534/PF2727/M3733, F719/M3636/IMS-JST003305/V1029/IMS-JST00305, P157/PF2771, L929/PF2605/M3643, L132.1/L132/L132.2/PF1437, YSC0001297/F1209/M3654/V1990, Y2888, P158/PF2706, PF2614/F1089/M3649/V1597, M3771/CTS12632/PF2775, M3673/CTS1911/PF2649, M3729/CTS9317/PF1767, PF2748/M3751, YSC0001308/PF2709/F2587/M3719, PF2736/F3111/M3740, PF2598, PF2600, PF2758/F3335/M3754, PF2731/F2985/M3736/V3919, L470/PF2730/M3735/V3900, PF2635/M9059, M3720/CTS8014/PF2711, PF2737/F3136/M3741, PF2592, M3712/CTS6542/PF2699, PF2616/F1149/M3653, Y1807/FGC6229, M235/PF2665/PAGE80/PAGES00080, M3706/CTS5948/PF1695, PF2639, M3689/CTS3654/PF2667, PF2597, PF2729/F2964, PF2742/F3254/M3746, M213/P137/PF2673/PAGES00038/PAGE38, CTS10213/PF2733/M3738, PF2770/M3767, F3556/M3765/PF1914, P161/PF2719, Y1822, M3640/CTS540/PF1506, PF2621/F1320/M3657, CTS5264/PF2684, PF2643, CTS2220/PF2656/M3679/MF624945, PF2744/M3748, PF2634/M9058, PF2688/F2142, PF2593, PF2609/M3645, PF2628/M3662, PF2772/F3616/M3769, PF2749/M3752, Y1809/FGC2046, CTS6135/PF2693/M3708, L1074/CTS4267/PF2674/M3694, PF2626/F1416/M3660, M3682/CTS2480/PF2659, PF2752, Y1805/FGC2055, CTS3868/PF2669, PF2590/V205/M3638, PF2769/M3764, PF2653/F1714/M3677, M3773/CTS12673, PF2596, P139/PF2698, PF2740/M3744, M89/PF2746, M3724/CTS8638, PF2627/M3661, CTS3536/PF2666/M3688, PF2588/M3635/CTS71, CTS12138/PF2774/M3770, PF2743/M3747, PF2594, PF2695/M3710/F3947, P187/PF2632, P316/PF2696, F3692/M3650, PF2658/F1753/M3681, PF2700/F2402/M3714, PF2722/F2837/M3726/V3477, M3713/CTS6843/PF1720, PF2608, PF2718/F2710/M3723, YSC0001295/PF2610/F1046/M3646/V1355, P142/PF2604, PF2631/M3665, CTS7002/PF2701/M3715, CTS4557/PF2679/M3698, CTS1468/PF2607/M3644, M3696/CTS4443/PF2677, CTS5432/PF2687/M3702, M3721/CTS8467/PF2715, M3687/CTS3195/PF2664, PF2619/F1285/M3655/V2194, F3512/PF1911, Y1803/FGC2053, M3728/CTS9280/PF2724, Y1463/FGC7686, P163/PF2686, L851/CTS11821/PF2767/M3762, F773, PF2630/M3664, CTS11150/PF2761/M3758, PF2690/F2155/M3704, P160/PF2618, P140/PF2703, Y1820/FGC2062, PF2613, M3755/L508/PF2759/FGC2052, CTS11819/PF2766/M3761, PF2646, PF2738/M3742, P134/PF2606, CTS3944/PF2670/M3691, M3697/CTS4470/PF2678, PF2611/M3647 P166/PF2702, Y1808, CTS2097/PF2654/M3678, CTS10290/PF2735/M3739, Y1812, L313/PF1426/M3651/V1644, P14/PF2704, PF2750/M3753, F3561/M3766, P141/PF2602, M3718/CTS7878, P136/PF2762, PF2685/F2075, L543/PF2663/M3686/V2513, P146/PF2623, L350/PF2692/M3707, Y1799/FGC2061, CTS1932/PF2650/M3674, PF2716/F2688/M3722, PF2629/M3663, PF2612/M3648, P149/PF2720, L498/PF2707/M3717, M3731/CTS9418/PF2726, M3756/CTS10983/PF2760, PF2713, PF2660/F1767/M3683, CTS2041/PF2652/M3676, PF2599, Y1813, P133/PF2636.
https://discover.familytreedna.com/y-dna/F-M89/story [30]
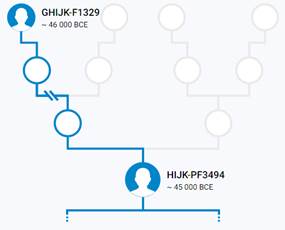
Предок (сопредок) GHIJK-F1329 (+ женщина): у него на Y-хромосоме в позиции 8720990 произошла необратимая однонуклеотидная мутация С > Т. По данным YFull это событие произошло примерно 48800 лет назад в интервале 51400 <–> 46300 лет назад (formed CI 95%). Филогенетические сопредки узла GHIJK-F1329 – это приблизительно 2 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: YSC0001299/PF2622/F1329/M3658/V2308, M3684/CTS2569/PF2661.
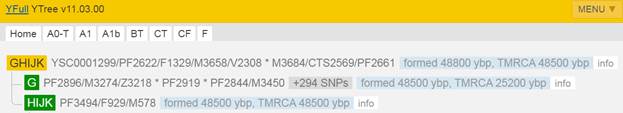
https://www.yfull.com/tree/GHIJK/ [40]
https://discover.familytreedna.com/y-dna/GHIJK-F1329/story [30]
Предок (сопредок) HIJK-PF3494 (+ женщина): у него на Y-хромосоме в позиции 7334662 произошла необратимая однонуклеотидная мутация С > Т. По данным YFull это событие произошло примерно 48500 лет назад в интервале 50900 <–> 46200 лет назад (formed CI 95%). Синониминые названия для SNP PF3494 – F929 и M578.
https://discover.familytreedna.com/y-dna/HIJK-PF3494/story [30]
Предок (сопредок) IJK-L15 (+ женщина): у него на Y-хромосоме в позиции 6885478 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 48500 лет назад в интервале 50900 <–> 46200 лет назад (formed CI 95%). Филогенетические сопредки узла IJK-L15 – это приблизительно 6 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: L16/M522/S138/PF3493, YSC0001319/PF3497/M2684/V1438, L15/M523/S137/PF3492/Z4413, PF3495/F3689/M2682/Y2571/V1295, PF3500/M2696, YSC0001318/PF3496/M2683.
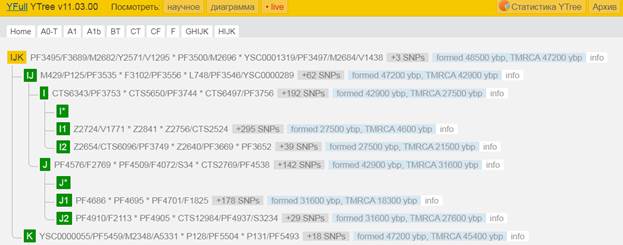
https://www.yfull.com/tree/IJK/ [40]
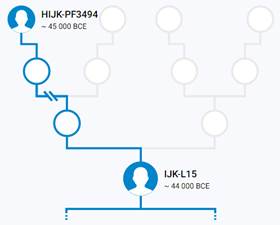
https://discover.familytreedna.com/y-dna/IJK-L15/story [30]
Предок (сопредок) K-M9 (+ женщина): у него на Y-хромосоме в позиции 19568371 произошла необратимая однонуклеотидная мутация С > G. По данным YFull это событие произошло примерно 47200 лет назад в интервале 49700 <–> 44600 лет назад (formed CI 95%). Филогенетические сопредки узла K-M9 – это приблизительно 21 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: PF5507/M2697, PF5469/V104, CTS10976/PF5509/M2698, PF5488/M2351, L469/PF5499/M2689, PF5490/F1765/M2685, PF5495/F2006/M2688/V3169, YSC0000055/PF5459/M2348/A5331, M2352/CTS2071/PF5489, PF5503/F3026/M2694/V4038, Y440/FGC221, L819/CTS4265/PF5494/M2686, PF5470, PF5500/F2548/M2692, YSC0000222/PF5505/L1346/M2695, CTS9278/PF5501/M2693, Y397, M9/PF5506/TY3327, P132/PF5480, P128/PF5504, P131/PF5493.
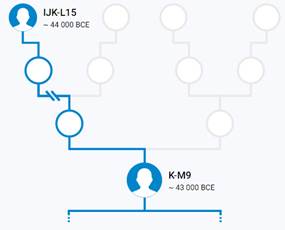
https://discover.familytreedna.com/y-dna/K-M9/story [30]
Предок (сопредок) K2-M526 (+ женщина): у него на Y-хромосоме в позиции 21389038 произошла необратимая однонуклеотидная мутация А > С. По данным YFull это событие произошло примерно 45400 лет назад в интервале 49600 <–> 41400 лет назад (formed CI 95%). Синониминое название для SNP M526 – PF5979.
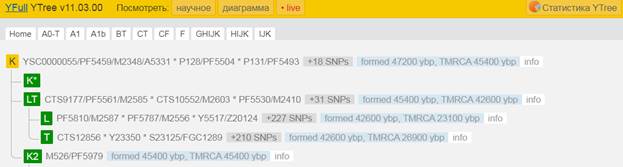
https://www.yfull.com/tree/K/ [40]
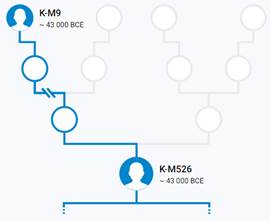
https://discover.familytreedna.com/y-dna/K-M526/story [30]
Предок (сопредок) K2b-YSC0000186 (+ женщина): у него на Y-хромосоме в позиции 14110681 произошла необратимая однонуклеотидная мутация С > Т. По данным YFull это событие произошло примерно 45400 лет назад в интервале 49600 <–> 41400 лет назад (formed CI 95%). Филогенетические сопредки узла K2b-V1651 – это приблизительно 5 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: MF44733/M1221/YSC0000186/PF5911/P331, PF5852, PF5969, L405/PF5990, M1205/CTS2019.
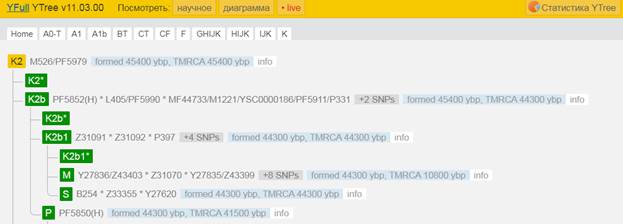
https://www.yfull.com/tree/K2/ [40]
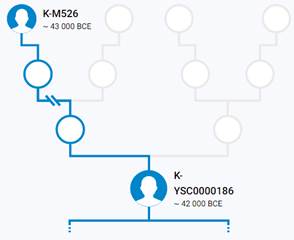
https://discover.familytreedna.com/y-dna/K-YSC0000186/story [30]
Предок (сопредок) P-PF5850 (+ женщина): у него на Y-хромосоме в позиции 4853930 произошла необратимая однонуклеотидная мутация Т > А. По данным YFull это событие произошло примерно 44300 лет назад в интервале 45400 <–> 43000 лет назад (formed CI 95%).
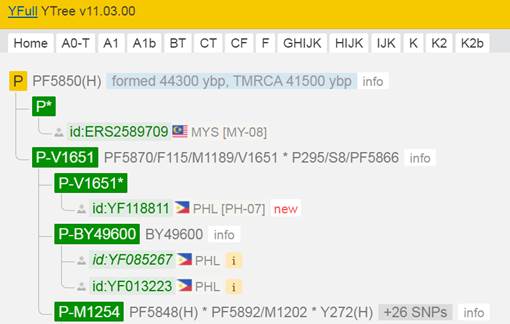
https://www.yfull.com/tree/P/ [40]
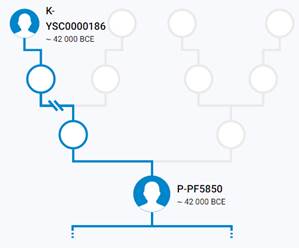
https://discover.familytreedna.com/y-dna/P-PF5850/story [30]
Предок (сопредок) P-V1651 (+ женщина): у него на Y-хромосоме в позиции 8266663 произошла необратимая однонуклеотидная мутация Т > А. В YFull YTree время появления данного субклада не указано, но, учитывая тот факт, что он следующий нисходящий после P-PF5850, то это событие произошло примерно тогда же, а именно 44300 лет назад в интервале 45400 <–> 43000 лет назад (formed CI 95%). Филогенетические сопредки узла P-V1651 – это приблизительно 2 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: PF5870/F115/M1189/V1651, P295/S8/PF5866.
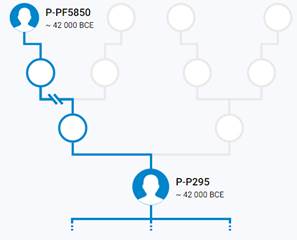
https://discover.familytreedna.com/y-dna/P-P295/story [30]
Предок (сопредок) P-M1254 (+ женщина): у него на Y-хромосоме в позиции 18960100 произошла необратимая однонуклеотидная мутация А > Т. В YFull YTree время появления данного субклада не указано, но, учитывая тот факт, что он следующий нисходящий после P-PF5850, то это событие произошло примерно тогда же, а именно 44300 лет назад в интервале 45400 <–> 43000 лет назад (formed CI 95%). Филогенетические сопредки узла P-M1254 – это приблизительно 29 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: Y448/FGC216, Y450, Y451, CTS3316/M1209, Y446, Y463/Z3131, M1270/CTS11173/PF5974, Y444, Y462/FGC212, Y267, Y458/FGC217, Y455/FGC213, PF5848, Y456/FGC286, CTS5418/PF5912/M1222, PF5853, PF5892/M1202, PF5461, PF5468, Y466, PF5968/M1266, Y272, Y1816, Y454, PF5935, PF5993/Z1244, PF6062/M1254, Y447/FGC211, Y503_2.
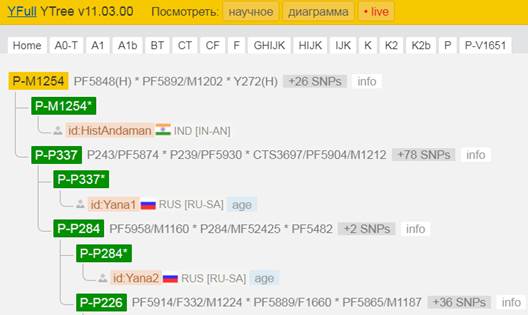
https://www.yfull.com/tree/P-M1254/ [40]
Предок (сопредок) P-P337 (+ женщина): у него на Y-хромосоме в позиции 12786160 произошла необратимая однонуклеотидная мутация А > G. В YFull YTree время появления данного субклада не указано. В FTDNA после субклада P-P295 сразу следует P-M45/PF5962, который филоэквивалентен P-P337, однако P-M45/PF5962 отсутствует в Y-отчёте. Филогенетические сопредки узла P-P337 – это приблизительно 81 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: CTS3446/PF5902/M1211, PF5965/M1265, M1216/YSC0000176/PF5908/V2979, CTS7481/PF5926/M1234, PF5465, Y45/M1208, P27.1_1/P207/P27.2_1/P27_1, PF5471, YSC0001257/CTS1907/PF5894/M1204, V231/PF5862/F91, M1235/CTS7604/PF5928, M1186/YSC0000279/PF5864, CTS3509/PF5903/M1103, PF6066/M1258, CTS7244/PF5924/M1233, PF5855, L1185/CTS9162/PF5937/M1241, CTS12299/PF5987, PF5981/F671/M1274, Y474/M1191, PF5975/F640/M1271/MF53797, PF5881/F180/M1196, PF5952/M1255, PF5861/F83/M1185/V1195/MF37664, PF5980/F653, L471/PF5989, M1250/CTS10085/PF5948, L779/PF5907/YSC0000251/V2974, PF5956/M1259, M1218/CTS4944/PF5909/V3240, L821/PF5857/F29/M5579, M1228/CTS5884/PF5917, PF5849, Y1610, PF5887, F313/M1219, PF5885/F212/M1198, L721/PF6020, PF5991/F4/M1183, Y503_1, PF5483, PF5846/M1184/CTS216, MF49095/F506/PF5940/M1243/YSC0000966/V3529, PF5878/M1194/MF14680, M1269/CTS10859, M1237/CTS8356/PF5931, PF5869/M1188, PF5959/M1261, PF5845/CTS196/V1079, PF5867, PF5888/V5158, CTS6948/PF5922/M1231/MF46647, PF5999, PF5978/F647/M1273, V607_1/Y483_1, CTS10348/PF5950/M1252, Y507/M1267, PF5851, L768/PF5976/YSC0000274, CTS8626/PF5934/M1239/MF48306, PF5876/M1193, M1246/YSC0000270/PF5943/V3732, M1220/CTS5340/PF5910/MF15401, PF5901/F1857/PAGES00083/PAGE83/P337, CTS3697/PF5904/M1212, CTS5808/PF5915/M1226, PF5871/M1190, M1249/CTS10081/PF5947, PF5982, PF5882, PF5916/F344/M1227, PF5886/M1199, M1238/CTS8473/PF5933, PF5464, FGC285, P244/PF5896/P244.1/P244.2, P237/PF5873, P281/PF5941, P239/PF5930, P228/PF5927, P243/PF5874.

https://discover.familytreedna.com/y-dna/P-M45/story [30]
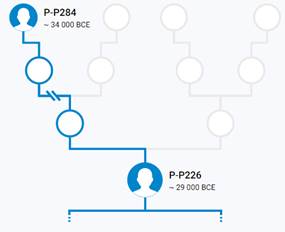
Предок (сопредок) P-P284 (+ женщина): у него на Y-хромосоме в позиции 19910454 произошла необратимая однонуклеотидная мутация С > G. В YFull YTree время появления данного субклада не указано. Филогенетические сопредки узла P-P284 – это приблизительно 5 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: PF5958/M1160, PF5482, PF5854, P284, PF5883/M1197.
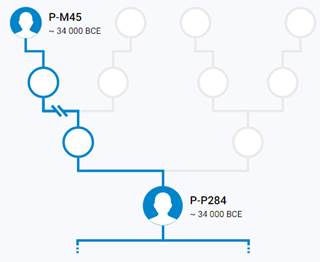
https://discover.familytreedna.com/y-dna/P-P284/story [30]
Предок (сопредок) P-P226 (+ женщина): у него на Y-хромосоме в позиции 8977339 произошла необратимая однонуклеотидная мутация С > Т. В YFull YTree время появления данного субклада не указано. Филогенетические сопредки узла P-P226 – это приблизительно 39 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: PF5487/M1201, M1109/CTS4437, PF5880/M1195, CTS3736/PF5905/M1213, PF5955/M1257, PF5954/M1256/MF51014, P235/PF5946, CTS12028/PF5977/M1272, M1264/YSC0000227/F597, CTS7886/PF5929/M1236, CTS3135/PF5898/M1206, P283/PF5966, PF5951/F556, M1253/CTS10454, PF5964/M1263, PF5476, CTS3813/PF5491/M1215, PF5872/M1192/V1809.1, M1232/CTS7194/PF5923, M1240/YSC0000205/PF5936, CTS3358/M1210/PF5899/PF5900, PF5994, M1149/CTS10168/PF6061, L82/PF5972, PF5971, YSC0001285/CTS5673/PF5497/M1225, PF5865/M1187, PF5985, L781/PF5875/YSC0000255, Y1403/FGC83, PF5914/F332/M1224, PF5920/F359/M1229, L536/PF5860, PF5889/F1660, PF5945/F524/M1248, PF5473, P230/PF5925, M74/N12/PF5963/MF52086, P226/PF5879.
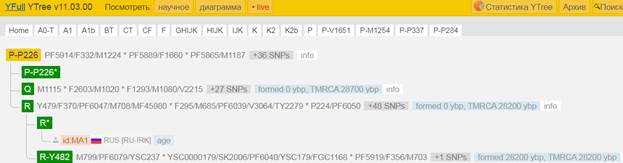
https://www.yfull.com/tree/P-P226/ [40]
https://discover.familytreedna.com/y-dna/P-P226/story [30]
Предок (сопредок) R-M207 (+ женщина): у него на Y-хромосоме в позиции 13470103 произошла необратимая однонуклеотидная мутация A > G. В YFull YTree время появления данного субклада не указано, приведено лишь TMRCA – 28200 лет назад в интервале 30500 <–> 25900 лет назад (formed CI 95%). Филогенетические сопредки узла R-M207 – это приблизительно 51 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: Y472/F47/M607/PF6014/S9, M732/CTS8311/PF6055, M696/CTS5815/PF6044, L248.1/L248.2/PF6045/L248/M705/M705.3, V3272, CTS9200/PF5938/V3466, Y480, PF5868/M628, L760/PF5877/YSC0000286/M642, Y471, PF5953/M764, L747/PF5918/YSC0000287/M702, Y437/FGC207, Y479/F370/PF6047/M708/MF45980, F295/M685/PF6039/V3064/TY2279, Y441, YSC0001265/CTS3229/PF6036/M672/V2573, F675/PF6084, CTS7876/PF6052, PF5992/M600/CTS207, M788/CTS10663/PF6075, CTS2913/PF6034/M667, PF6013/F33/M603, A18557/M795/CTS11075/PF6078, Y296/M651/PF6024, PF6002, M741/CTS9005/PF6058, F154/M636/PF6021/V2252, F652/M805/PF6082, Y453/FGC202, P227, Y460/FGC204, CTS7880/PF6053/M725, YSC0000233/PF6077/L1347/M792, PF6016/F63/M614, Y442/FGC205, CTS3622/PF6037, YSC0000232/M789/L1225/PF6076, Y506/M760/PF6063, F82/M620/V1194/MF37663, CTS2426/PF6033/M661, Y469, Y457/FGC208, P280/PF6068, P224/PF6050, M207/UTY2/PF6038/PAGES00037/PAGE37, P285/PF6059/FGC201, P232, YSC0000201/PF6057/M734/S4, Y125018, P229/PF6019.
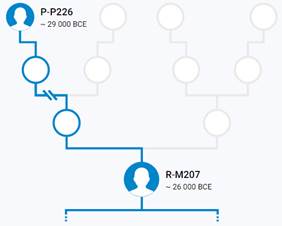
https://discover.familytreedna.com/y-dna/R-M207/story [30]
Предок (сопредок) R-Y482 (+ женщина): у него на Y-хромосоме в позиции 15905648 произошла необратимая однонуклеотидная мутация G > Т. По данным YFull это событие произошло примерно 28200 лет назад в интервале 30500 <–> 25900 лет назад (formed CI 95%). Филогенетические сопредки узла R-Y482 – это приблизительно 4 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: Y482/PF6056/F459, YSC0000179/SK2006/PF6040/YSC179/FGC1168, PF5919/F356/M703, M799/PF6079/YSC237.
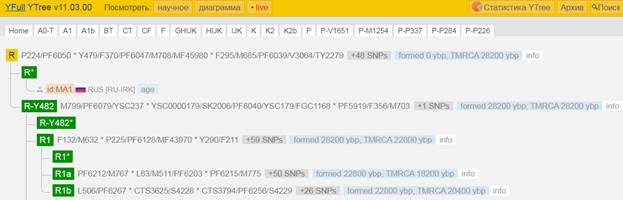
https://www.yfull.com/tree/R/ [40]
Предок (сопредок) R1-M173 (+ женщина): у него на Y-хромосоме в позиции 12914512 произошла необратимая однонуклеотидная мутация А > C[26]. По данным YFull это событие произошло примерно 28200 лет назад в интервале 30500 <–> 25900 лет назад (formed CI 95%). Филогенетические сопредки узла R1-M173 – это приблизительно 62 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: PF6120, M710/YSC0000192/PF6132, PF6073, Y481/M716, PF6011/FGC193, Y305/PF6031/FGC189, M717/CTS7122/PF6135, PF6118/M640, M654/CTS1913/PF6032, PF6110, Y449, M306/S1/PF6147/MF53060, Y464/PF6008/FGC218, Y400, YSC0001281/CTS4862/PF6042/M691, PF5859/M611/CTS916/Z2133, CTS3321/PF6125/M673, M694/CTS5611/PF6130, L875/PF6131/YSC0000288/M706, CTS3123/PF6124/M670, P233/PF6142, Y459, M748/YSC0000207, Y477/PF6121/F245/M659, Y512, M781/PF6145, Y452/FGC203, YSC0000230/L1352/M785/BZ3050, CTS2680, M813/CTS12618/PF6089, P245/PF6117, Y436, Y513, PF6007, F132/M632, P294/PF6112/FGC13894, M643, FGC190, Y470, PF6119, PF6133/F378/M711, P286/PF6136, M682/CTS4075/PF6127, M714/CTS7066/PF6049, P242/PF6113, P238/PF6115, PF6116/F102/M625/V1478, Y467/FGC194, FGC206, M663/CTS2565/PF6122, P236/PF6137, P225/PF6128, M812/CTS12546/PF6088, PF6146/FGC465, CTS2908/PF6123/M666, Y465/FGC198, M730/CTS8116/PF6138, Y290/F211, PF6111/M612/CTS997, P234/PF6141, M173/P241/PF6126/PAGES00029/PAGE29, Y125013.
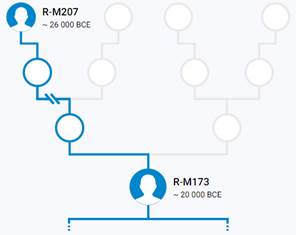
https://discover.familytreedna.com/y-dna/R-M173/story [30]
Предок (сопредок) R1а-M420 (+ женщина): у него на Y-хромосоме в позиции 21311315 произошла необратимая однонуклеотидная мутация Т > А. По данным YFull это событие произошло примерно 22800 лет назад в интервале 25100 <–> 20500 лет назад (formed CI 95%). Филогенетические сопредки узла R1а-M420 – это приблизительно 53 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: PF7527/F2948/M752/V3820, CTS5164, Y209/FGC32015, CTS6918/PF6196, CTS12321, CTS8851/M740, CTS11734/PF6226/M800, CTS9515/M744, CTS5936/PF6192/M698, L145/M449/PF6175, PF6179/F1769/M662, PF6153/F886, CTS4509/M687, L63/M511/PF6203, PF6160/F1088/M629, CTS7559, CTS3877/PF6184, CTS11530, CTS9596/PF6205/M745/V3655, PF6233/F3570, Y216/M784, PF6163, Y218, PF6213/M768, M644, CTS2907/M665, PF6212/M767, CTS11148/M796, PF7534/F3466/M803, CTS8008/M726, CTS12746/M815, Y215/FGC32014, PF6222/F3364/M794, CTS10627/M786, L62/M513/PF6200, L566/PF7512, L146/M420/PF6229, PF6215/M775, Y217/FGC32438, CTS903/PF6154/M610, CTS5273/PF6190, L457/PF6191, F928/M616, CTS9667, Y194, Y1404/FGC89, Y195, Y1424/FGC92, Y190, CTS12639/PF7535/M814, Y212/M783, PF6189, PF7516/M641.
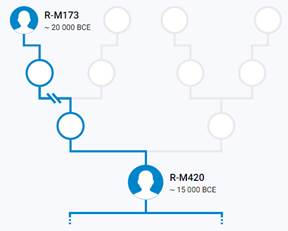
https://discover.familytreedna.com/y-dna/R-M420/story [30]
Предок (сопредок) R-M459 (+ женщина): у него на Y-хромосоме в позиции 7038033 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 18200 лет назад в интервале 20100 <–> 16400 лет назад (formed CI 95%). Филогенетические сопредки узла R-M459 – это приблизительно 54 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: Y1975/FGC2548, CTS11411/M798, CTS9739/M749, CTS10847/PF6221/M791, PF7531, PF6159/M626, F1224/V2035, CTS10042/PF6207/M754, PF6194/F2215/M701, PF6172, CTS5287, PF7514/M623/V1412, PF6230/F3494/M804, PF6170/M650, PF6204/F2901/M742/V3610, M459/PF6235, CTS3984/PF6185/M681/V2878, F937, CTS3548/M678/V2747, L122/M448/PF6237, PF6156/F947/M617, PF7522/F2328/M709, CTS7500/PF6199/M722, L120/M516/PF6236, PF7542, Y214/FGC32011, CTS11633, PF6214/M769, F3197/M774, PF6168/F1545/V5016, CTS3943/M680, CTS2132/PF6176/M658, F3564/M5824/F3564.2/M5824.2, Y172, PAGE65.2!/SRY1532.2!/SRY10831.2!/PF6234!/Page65.1/SRY10831.2/PAGE65.2/SRY1532.2, CTS11853/PF6227, PF6164/F1157/M631/V1855, PF6211/M766, PF6220, Y1270, CTS501/PF6152/M604, Y187, CTS1963, Y1420, CTS11706, Y173, Y1425/FGC90, Y183, PF6151, CTS2443/PF6178, FGC85, M602, Y191, CTS836/M609.
https://www.yfull.com/tree/R-M459/ [40]
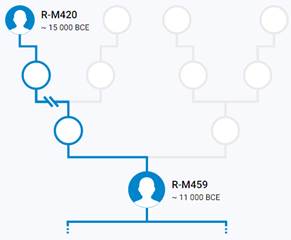
https://discover.familytreedna.com/y-dna/R-M459/story [30]
Предок (сопредок) R-M735 (+ женщина): у него на Y-хромосоме в позиции 16009389 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 15200 лет назад в интервале 17600 <–> 12900 лет назад (formed CI 95%). Филогенетические сопредки узла R-M735 – это приблизительно 6 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: CTS3527/M676, CTS5437/M693, PF7528/M770, CTS4812/PF6187/M690, CTS8637/M736, CTS8636/M735.
Предок (сопредок) R-M198 (+ женщина): у него на Y-хромосоме в позиции 12918840 произошла необратимая однонуклеотидная мутация С > Т. По данным YFull это событие произошло примерно 14000 лет назад в интервале 15600 <–> 12400 лет назад (formed CI 95%). Филогенетические сопредки узла R-M198 – это приблизительно 57 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: CTS262/M601, Y210/FGC32013, CTS9690/M747, CTS9779, Y220/M807, F4138, L449/PF6223, Y2363/M12464, CTS11184/PF6224/M797, M512/PF6239, CTS570, CTS3551/PF6183, F4099/M633/V1902, L168, PF6181/F1808/M668, Y206/M758, F989/M619/V1190, CTS8797/M738, CTS2891/M664, M647, Y170, M514/PF6240, Y221/M808, CTS3534/PF7518/M677, F3194/M773, CTS11720, Y205/FGC32012, M515, CTS4465/PF7519/M686, PF6216/M779, CTS3230, Y196/FGC36392, PF6201/F2684/M733, F1050/M622/V1380, F3337, M198/PF6238, F3185/M771, CTS1619/PF6173/M653, Y185, PF6167/M646, CTS8710/M737, CTS7072/M715, CTS3004/M669, Y188, Y179, Y186, CTS8073/M728, Y189, CTS8090/M729, Y184/Z10068, CTS7690/M724, Y192, Y207/M765, CTS9496/PF7526/M743, Y174, CTS3161/PF6182/M671, CTS8825/M739.
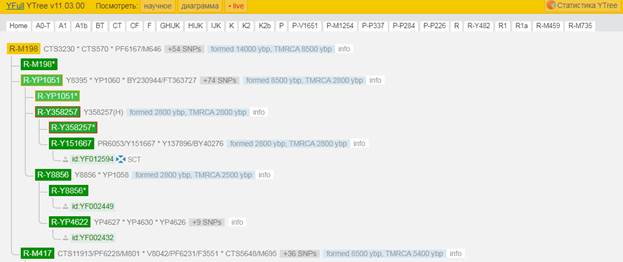
https://www.yfull.com/tree/R-M198/ [40]
https://discover.familytreedna.com/y-dna/R-M198/story [30]
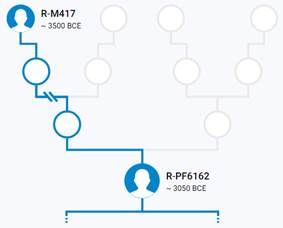
Предок (сопредок) R-M417 (+ женщина): у него на Y-хромосоме в позиции 8665694 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 8700 лет назад в интервале 10000 <–> 7500 лет назад (formed CI 95%). Филогенетические сопредки узла R-M417 – это приблизительно 39 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: CTS7278/PF7524/M721, PF6210/M757, CTS11913/PF6228/M801, CTS1340/PF6157/M618, Y1974/FGC2547, V8042/PF6231/F3551, CTS7191/PF6198/M719, CTS10993/M793, CTS5069, PF7530, CTS4259/PF6186/M683, PF6195/F2234/M704, PF7540/M759, PAGE7/PAGES00007, CTS5648/M695, PF7532/F3398, M630/V1720, CTS5423/M692, M417, Y1976/FGC2550, CTS10080/PF6208/M755, CTS9510/V3625, PF6169/M649, F3166/M763, M627, F3159/M761, PF6218/M782, CTS12941/M817, PF6165/M637, CTS5979/PF6193/M700, Y219/M806, F2957/M753/V3842, CTS6423, FGC87, Y176, CTS6544/M712, CTS1924/PF6174/M655, Y171, Y181.
https://discover.familytreedna.com/y-dna/R-M417/story [30]
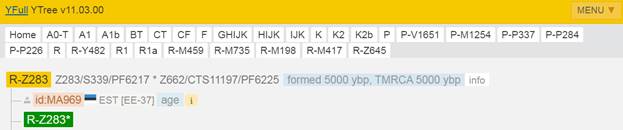
Предок (сопредок) R-Z645 (+ женщина): у него на Y-хромосоме в позиции 8377004 произошла необратимая однонуклеотидная мутация С > Т. По данным YFull это событие произошло примерно 5400 лет назад в интервале 6000 <–> 4700 лет назад (formed CI 95%). Филогенетические сопредки узла R-Z645 – это приблизительно 8 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: Z647/S441/PF6158, Z651/F3044/V4100, Z649/CTS5508, Z648/CTS12010/PF7533/M802, Z650/CTS9754/PF6206/M750/V3726, Z645/S224/PF6162/V1754, Z646/CTS6596/M713/S346, CTS12179/M811.
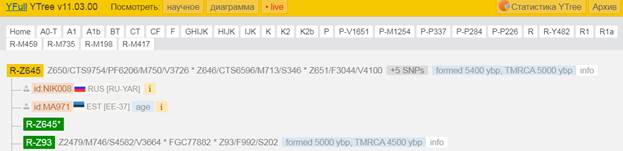
https://www.yfull.com/tree/R-Z645/ [40]
https://discover.familytreedna.com/y-dna/R-PF6162/story [30]
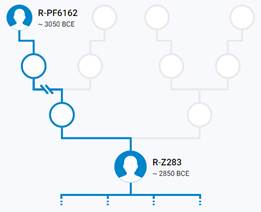
Предок (сопредок) R-Z283 (+ женщина): у него на Y-хромосоме в позиции 19814417 произошла необратимая однонуклеотидная мутация T > А. По данным YFull это событие произошло примерно 5000 лет назад в интервале 5400 <–> 4600 лет назад (formed CI 95%). Филогенетические сопредки узла R-Z283 – это приблизительно 2 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: Z283/S339/PF6217, Z662/CTS11197/PF6225.
https://www.yfull.com/tree/R-Z283/ [40]
https://discover.familytreedna.com/y-dna/R-Z283/story [30]
Предок (сопредок) R-Z282 (+ женщина): у него на Y-хромосоме в позиции 13476521 произошла необратимая однонуклеотидная мутация T > C. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%). Синониминые названия для SNP Z282 – S198 и V3055.
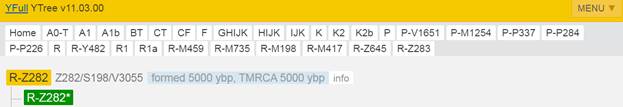
https://www.yfull.com/tree/R-Z282/ [40]
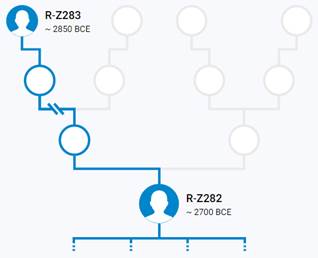
https://discover.familytreedna.com/y-dna/R-Z282/story [30]
Предок (сопредок) R-PF6155 (+ женщина): у него на Y-хромосоме в позиции 7387314 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%). Филогенетические сопредки узла R-PF6155 – это приблизительно 7 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: Y2459/FGC2603/M12404, Z2915, PF6161/Z2906/V1747, Z2912, Z2913, PF7525/Z2910, PF6155/Z2905/S4545.
https://www.yfull.com/tree/R-PF6155/ [40]
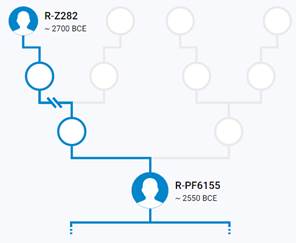
https://discover.familytreedna.com/y-dna/R-PF6155/story [30]
Предок (сопредок) R-M458 (+ женщина): у него на Y-хромосоме в позиции 22220317 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%). Филогенетические сопредки узла R-M458 – это приблизительно 4 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: M458/PF6241, PF6219/Z2911/S4549, PF6202/Z2909/S4555, Y2911/FGC2634/M12425.
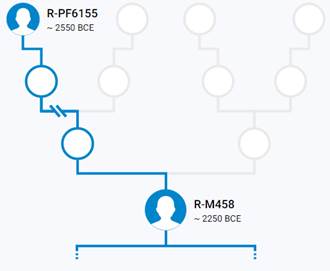
https://discover.familytreedna.com/y-dna/R-M458/story [30]
Предок (сопредок) R-PF7521 (+ женщина): у него на Y-хромосоме в позиции 13917476 произошла необратимая однонуклеотидная мутация A > C. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%). Синониминые названия для SNP PF7521 – Z2908 и S4567.
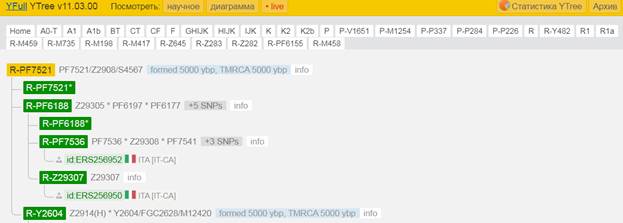
https://www.yfull.com/tree/R-PF7521/ [40]
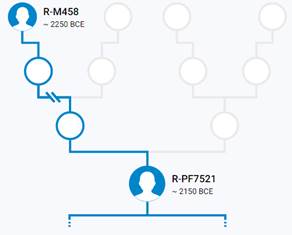
https://discover.familytreedna.com/y-dna/R-PF7521/story [30]
Предок (сопредок) R-Y2604 (+ женщина): у него на Y-хромосоме в позиции 15955572 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%). Филогенетические сопредки узла R-Y2604 – это приблизительно 2 потенциальных биологических мужских предка, которые поименованы следующим образом, включая синонимичные обозначения: Z2914, Y2604/FGC2628/M12420.
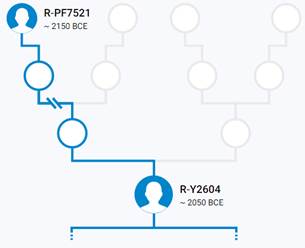
https://discover.familytreedna.com/y-dna/R-Y2604/story [30]
Предок (сопредок) R-FGC2608 (+ женщина): у него на Y-хромосоме в позиции 4311764 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%).
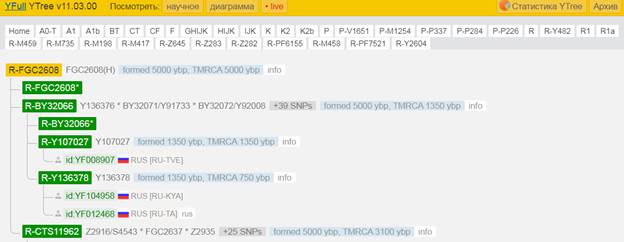
https://www.yfull.com/tree/R-FGC2608/ [40]
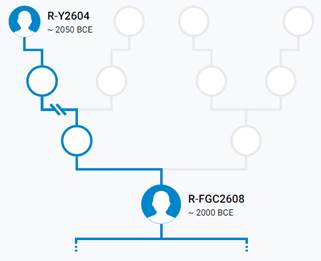
https://discover.familytreedna.com/y-dna/R-FGC2608/story [30]
Предок (сопредок) R-CTS11962 (+ женщина): у него на Y-хромосоме в позиции 21221843 произошла необратимая однонуклеотидная мутация C > T. По данным YFull это событие произошло примерно 5000 лет назад в интервале 6000 <–> 4000 лет назад (formed CI 95%). Синониминые названия для SNP YP417 – V3192, FGC20517, M12415. Филогенетические сопредки узла R-CTS11962 – это приблизительно 28 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: Z2947, Z2919/V1500, Z2932, Z2942, Y2462, Z2929/S4542, Z2933, Z2936, Z2934/S4557, Z2937, Z2930, Z2935, Z2948/V1632, FGC2637, Y2461, Z2943, Z2931, Z2939, Z2918, CTS11962/Z2953/Z2953.1/Z2953.2, Z2940, Z2928, Z2941, Z2944, Z2916/S4543, Y2460/FGC7770, Z2950, Z2921.
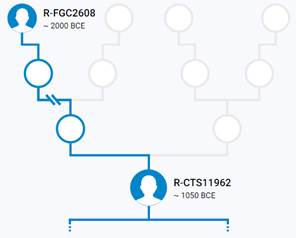
https://discover.familytreedna.com/y-dna/R-CTS11962/story [30]
Предок (сопредок) R-L1029 (+ женщина): у него на Y-хромосоме в позиции 15825380 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 3200 лет назад в интервале 3800 <–> 2600 лет назад (formed CI 95%). Синониминые названия для SNP YP417 – V3192, FGC20517, M12415. Филогенетические сопредки узла R-L1029 – это приблизительно 7 потенциальных биологических мужских предков, которые поименованы следующим образом, включая синонимичные обозначения: L1029/S4554, Z2951, Y5342/FGC2617, Z2922, Z2954/S4548, Z2920, Z2938/S4546.
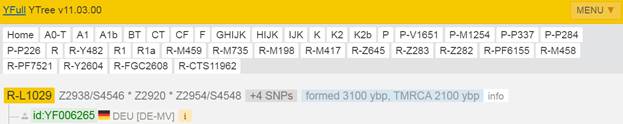
https://www.yfull.com/tree/R-L1029/ [40]
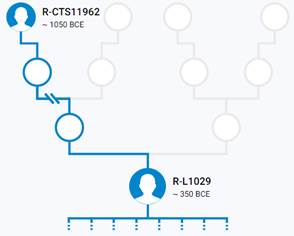
https://discover.familytreedna.com/y-dna/R-L1029/story [30]
Предок (сопредок) R-YP417 (+ женщина): у него на Y-хромосоме в позиции 13756598 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 2100 лет назад в интервале 2300 <–> 1900 лет назад (formed CI 95%). Синониминые названия для SNP YP417 – V3192, FGC20517, M12415.
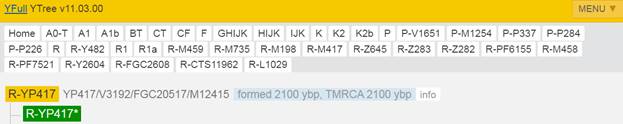
https://www.yfull.com/tree/R-YP417/ [40]
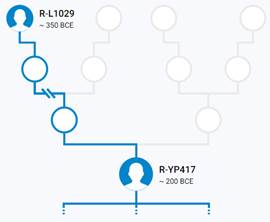
https://discover.familytreedna.com/y-dna/R-YP417/story [30]
Предок (сопредок) R-YP418 (+ женщина): у него на Y-хромосоме в позиции 7779691 произошла необратимая однонуклеотидная мутация A > T. По данным YFull это событие произошло примерно 2100 лет назад в интервале 2400 <–> 1800 лет назад (formed CI 95%). Синониминые названия для SNP YP418 – M12403 и V1330.
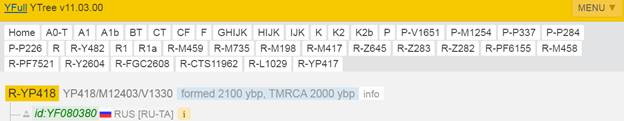
https://www.yfull.com/tree/R-YP418/ [40]
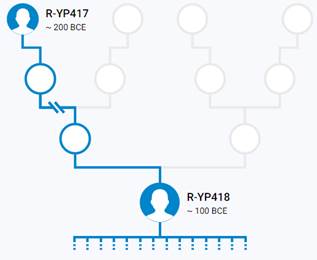
https://discover.familytreedna.com/y-dna/R-YP418/story [30]
Предок (сопредок) R-YP728 (+ женщина): у него на Y-хромосоме в позиции 7339125 произошла необратимая однонуклеотидная мутация G > C. По данным YFull это событие произошло примерно 2000 лет назад в интервале 2200 <–> 1850 лет назад (formed CI 95%).
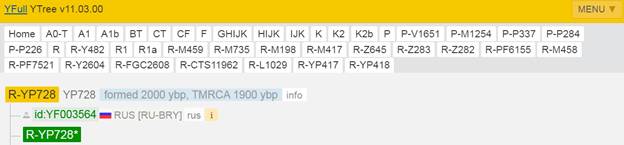
https://www.yfull.com/tree/R-YP728/ [40]
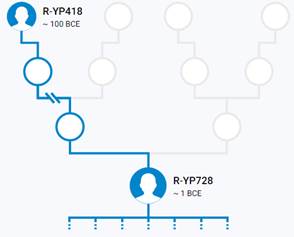
https://discover.familytreedna.com/y-dna/R-YP728/story [30]
Предок (сопредок) R-Y18348 (+ женщина): у него на Y-хромосоме в позиции 9264197 произошла необратимая однонуклеотидная мутация C > T. По данным YFull это событие произошло примерно 1900 лет назад в интервале 2200 <–> 1600 лет назад (formed CI 95%). Филогенетические сопредки узла R-Y18348 – это приблизительно 2 потенциальных биологических мужских предка, которые поименованы следующим образом: Y18348, Y18349.
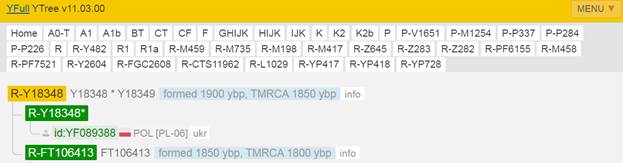
https://www.yfull.com/tree/R-Y18348/ [40]
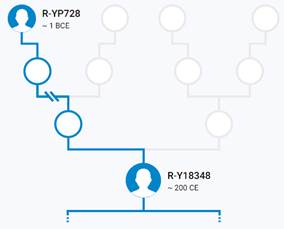
https://discover.familytreedna.com/y-dna/R-Y18348/story [30]
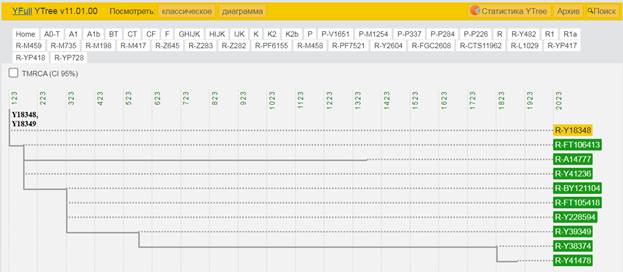
https://www.yfull.com/sc/tree/R-Y18348/ [40]
Предок (сопредок) R-FT106413 (+ женщина): у него на Y-хромосоме в позиции 26418953 произошла необратимая однонуклеотидная мутация A > G. По данным YFull это событие произошло примерно 1900 лет назад в интервале 2500 <–> 1350 лет назад (formed CI 95%).
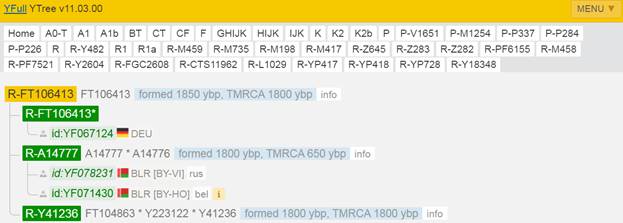
https://www.yfull.com/tree/R-FT106413/ [40]
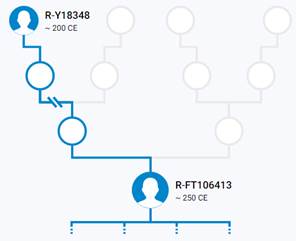
https://discover.familytreedna.com/y-dna/R-FT106413/story [30]
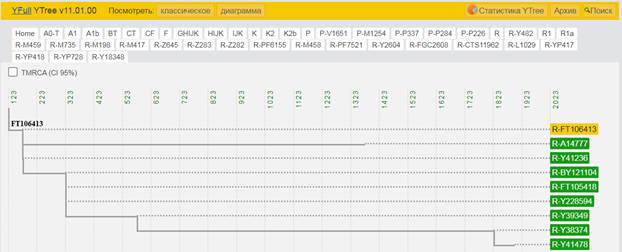
https://www.yfull.com/sc/tree/R-FT106413/ [40]
Предок (сопредок) R-Y41236 (+ женщина): у него на Y-хромосоме в позиции 13805982 произошла необратимая однонуклеотидная мутация G > A. По данным YFull это событие произошло примерно 1850 лет назад в интервале 2600 <–> 1200 лет назад (formed CI 95%). Филогенетические сопредки узла R-Y41236 – это приблизительно 3 потенциальных биологических мужских предка, которые поименованы следующим образом: Y41236, FT104863, Y223122.
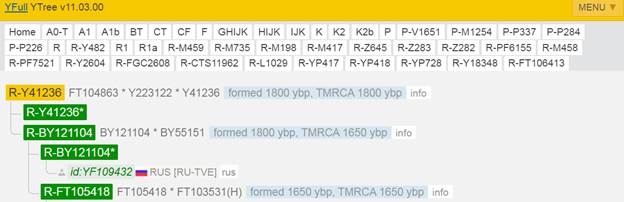
https://www.yfull.com/tree/R-Y41236/ [40]

https://www.yfull.com/sc/tree/R-Y41236/ [40]
Предок (сопредок) R-BY121104 (+ женщина): у него на Y-хромосоме в позиции 16127684 произошла необратимая однонуклеотидная мутация T > C. По данным YFull это событие произошло примерно 1850 лет назад в интервале 2600 <–> 1200 лет назад (formed CI 95%). Филогенетические сопредки узла R-BY121104 – это приблизительно 2 потенциальных биологических мужских предка, которые поименованы следующим образом: BY121104, BY55151.
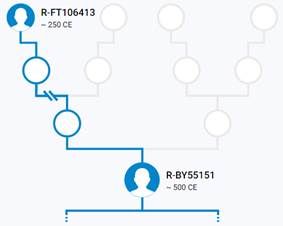
https://discover.familytreedna.com/y-dna/R-BY55151/story [30]
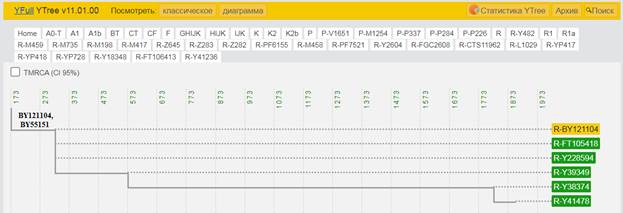
https://www.yfull.com/sc/tree/R-BY121104/ [40]
Предок (сопредок) R-FT105418 (+ женщина): у него на Y-хромосоме в позиции 14986412 произошла необратимая однонуклеотидная мутация А > T. По данным YFull это событие произошло примерно 1700 лет назад в интервале 2200 <–> 1200 лет назад (formed CI 95%). Филогенетические сопредки узла R-FT105418 – это приблизительно 2 потенциальных биологических мужских предка, которые поименованы следующим образом: FT105418, FT103531.
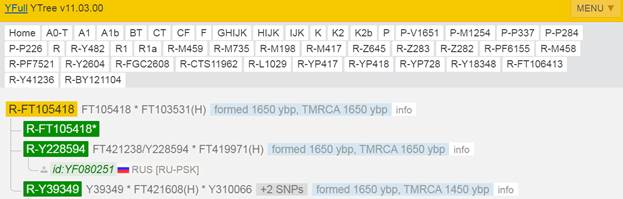
https://www.yfull.com/tree/R-FT105418/ [40]
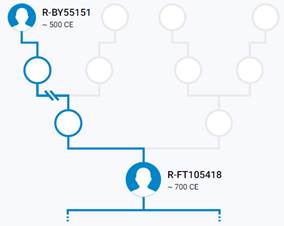
https://discover.familytreedna.com/y-dna/R-FT105418/story [30]
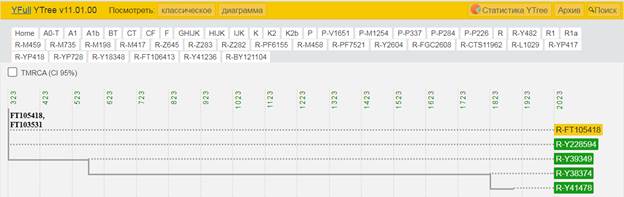
https://www.yfull.com/sc/tree/R-FT105418/ [40]
Предок (сопредок) R-Y39349 (+ женщина): у него на Y-хромосоме в позиции 8325641 произошла необратимая однонуклеотидная мутация А > T. По данным YFull это событие произошло примерно 1700 лет назад в интервале 2200 <–> 1200 лет назад (formed CI 95%). Филогенетические сопредки узла R-Y39349 – это приблизительно 5 потенциальных биологических мужских предков, которые поименованы следующим образом: Y39349, FT421608, Y43577, Y310066, Y263186.
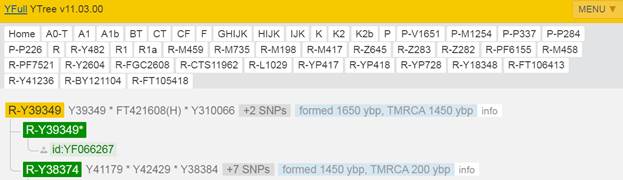
https://www.yfull.com/tree/R-Y39349/ [40]
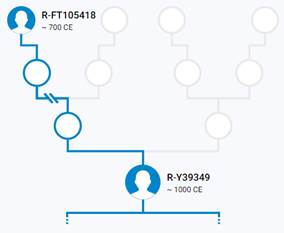
https://discover.familytreedna.com/y-dna/R-Y39349/story [30]
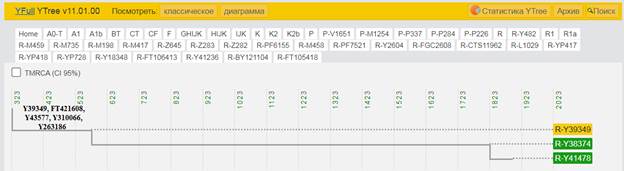
https://www.yfull.com/sc/tree/R-Y39349/ [40]
Предок (сопредок) R-Y38374 (+ женщина): у него на Y-хромосоме в позиции 2827785 произошла необратимая однонуклеотидная мутация G > T. По данным YFull это событие произошло примерно 1450 лет назад в интервале 2100 <–> 900 лет назад (formed CI 95%) и оно не относится к документально-реконструированной родословной Нилоговых. Филогенетические сопредки узла R-Y38374 – это приблизительно 10 потенциальных биологических мужских предков, которые поименованы следующим образом: Y43086, Y42429, FT171875, Y40155, FT171852, Y41179, FT171889, Y38384, FT171841, Y38374. Они является филоэквивалентными сопредками для R-Y38374, то есть приходятся на интервал почти в полторы тысячи лет, учитывая тот факт, что по принятой в YFull методике расчётов одна SNP-мутация возникает примерно раз в 144 года.
Сопредок R-Y38374 в качестве узлового выбран случайно: пока не будет определена последовательность восходящих и нисходящих снипов/сопредков – любой из десяти филоэквивалентов может стоять в основе ветвления линии. Важно то, что каждый из десяти – это реальный биологический мужской предок, у которого в Y-хромосоме однажды произошла необратимая SNP-мутация, унаследованная потомками подобно фамилии. Конечно, на протяжении 1500 лет, которые отведены для данного субклада, пришлось больше, чем десять прямых мужских предков: часть из них приходится на документально-реконструированную родословную (до конца XVI века), а большинство – либо безымянно для классической генеалогии, либо поименовано филогенетическим способом, а именно посредством индексов SNP (см.: [48],[49])[27].
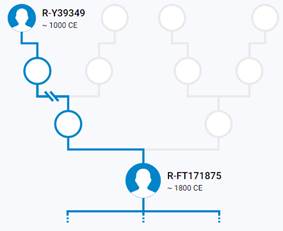
https://discover.familytreedna.com/y-dna/R-FT171875/story [30]
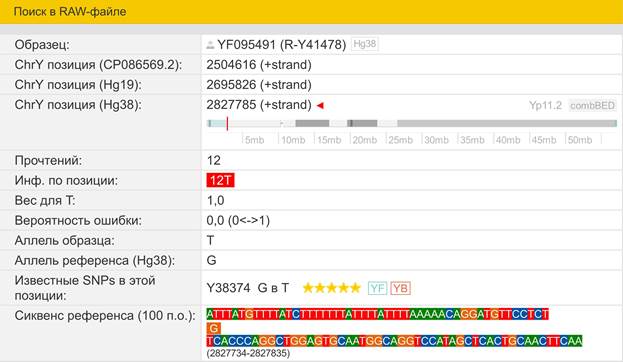
https://www.yfull.com/snp-to-showseq/95491/571547/ [32]
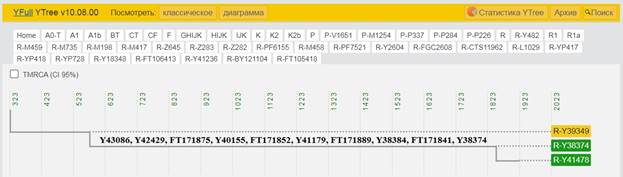
https://www.yfull.com/sc/tree/R-Y39349/ [40]
Предок (сопредок) R-Y41478 (+ женщина): у него на Y-хромосоме в позиции 14326593 произошла необратимая однонуклеотидная мутация C > T. По данным YFull это событие произошло примерно 150 лет назад, то есть относится к документально-реконструированной родословной Нилоговых второй половины XIX века. Поскольку эта мутация отсутствует у параллельной линии Нелоговых из Пензы, общий предок с которыми родился в 1847 году (Пётр Изосимович Нилогов), постольку она точно случилась у одного из наших прямых мужских предков – например, у Григория Петровича Нилогова, который родился в 1881 году. Его старший брат Антон Петрович Нилогов, родившийся около 1876 года, не имел этой мутации, иначе бы она передалась его прямым мужским потомкам, а у них она отрицательная. Не исключено, что SNP R-Y41478 могла впервые произойти у самого Петра Зосимовича, который в виде сперматозоида передал её своему позднему сыну Григорию (1881). Однако аналогичное предположение можно отнести и на счёт сына Григория – Фёдора (1909), а также на сына Фёдора – Михаила (1929). R-Y41478 выявлена положительной у сына Михаила – Сергея (1953), а, следовательно, к нему она перешла либо в сперматозоиде отца Михаила (то есть возникла до оплодотворения), либо случилась при оплодотворении у самого Сергея.
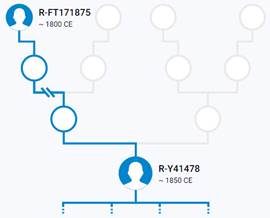
https://discover.familytreedna.com/y-dna/R-Y41478/story [30]
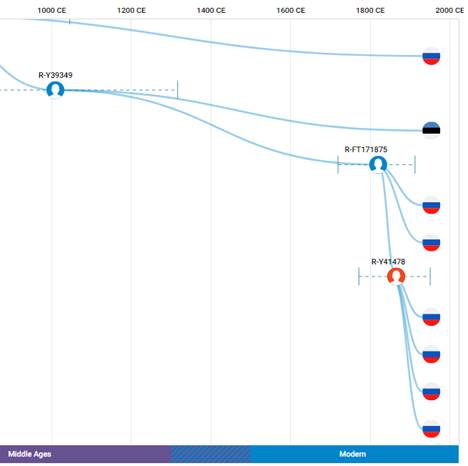
https://discover.familytreedna.com/y-dna/R-Y41478/tree [30]
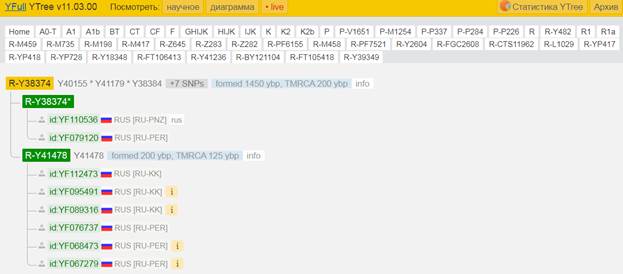
https://www.yfull.com/tree/R-Y38374/ [40]
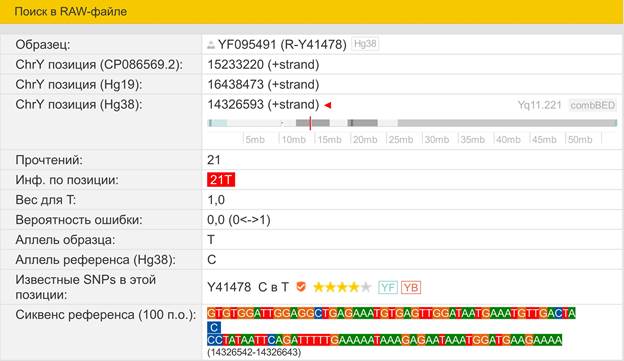
https://www.yfull.com/seq/38/95491/y/14326593/ [32]
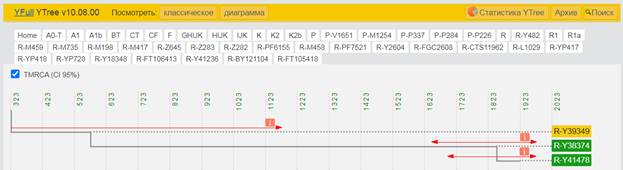
https://www.yfull.com/sc/tree/R-Y39349/ [40]
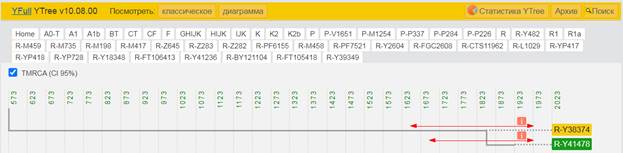
https://www.yfull.com/sc/tree/R-Y38374/ [40]

https://www.yfull.com/sc/tree/R-Y41478/ [40]
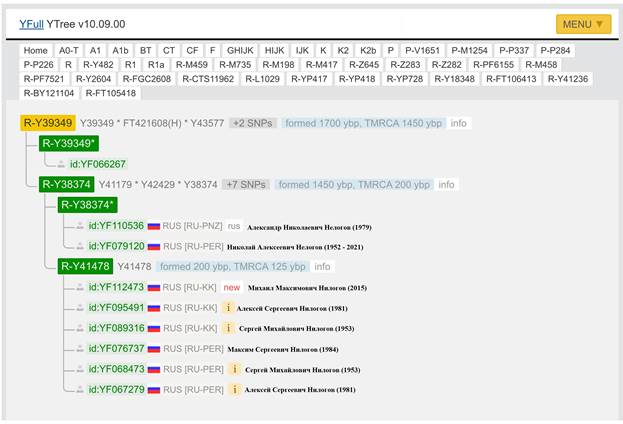
Атрибутированные доноры на YFull YTree: филогенетическая ветвь рода Нилоговых [40]
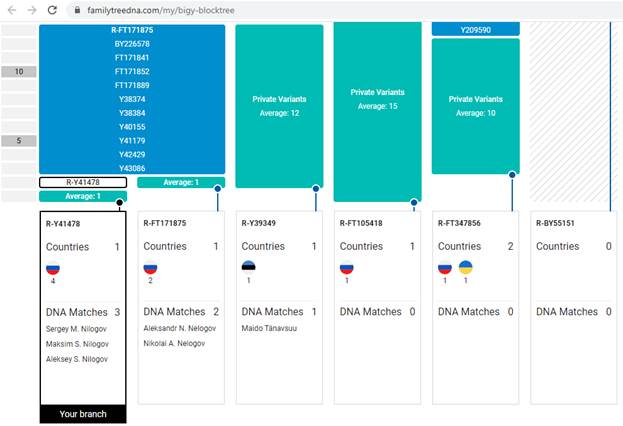
https://www.familytreedna.com/my/bigy-blocktree [58]
Таким образом, на примере генетико-генеалогической (генетеалогической) реконструкции патрилинии автора были показаны перспективы комплексного (междисциплинарного) исследования человеческого патрилинейного родства (см. также: [59],[60]). Проблема номинации наших далёких предков рассмотрена в аспекте лингвофилософской проблемы ономафикации как части патрофикации – общего дела по воскрешению отцов (Н. Ф. Фёдоров) [36]. Лингвогенетеалогический метод снипования как именования (антропонимофикации) от Y-хромосомного Адама через все узловые ветви (гаплогруппы/снипы) к документально подтверждённой фактологии классической генеалогии должен стать решением насущной проблемы по реконструкции популяционной и персональной генеалогии людей.
[1] Ср.: А. А. Клёсов: «Снип-мутации – это практически необратимые мутации в ДНК (от английского сокращения SNP, что означает «однонуклеотидные вариации»). Однажды образовавшись, они «застревают» в ДНК навсегда, за крайне редким исключением, когда в том же нуклеотиде, который ранее мутировал, прошла ещё одна мутация – либо в другой нуклеотид, либо возвращение в исходный. В книге «Ваша ДНК-генеалогия» был приведён пример, что один определённый нуклеотид Y-хромосомы у всех современных гоминидов (кроме макаки, которая относится к семейству мартышковых, хотя отряд тот же – приматы), выбранный для той иллюстрации, является тимином, и только у орангутана там цитозин. Иначе говоря, у большинства этих особей тимин сидит в том месте Y-хромосомы не менее 15–20 миллионов лет, и за это время из пяти «образцов» изменился лишь однажды. Скептик скажет, что он мог измениться в орангутанге за это время и сотню раз, и цитозин – просто последний в той серии изменений, но так не бывает. Уже накоплен большой массив экспериментальных данных, что мутации в ДНК происходят неупорядоченно, статистически, равновероятно по всем последовательностям, за исключением крайне редких случаев, когда отпадают или перестраиваются большие фрагменты ДНК, но здесь явно не тот случай.
Большая серия экспериментальных данных, полученная и перекрёстно проверенная разными коллективами исследователей, привела к средней величине константы скорости мутации в Y-хромосоме 0.82·10-9 на нуклеотид в год (более правильно – на пару нуклеотидных оснований в год). При такой скорости мутации за 5 миллионов лет, то есть до времени жизни общего предка шимпанзе и человека, в Y-хромосоме с её 58 миллионов пар оснований набежит 0.82·10-9 × 10·106 × 58·106 = 476 тысяч мутаций, что составит всего 0.8 % от 58 миллионов нуклеотидов. Здесь в расчётах положено 10 миллионов лет, потому что 5 миллионов лет прошло от общего предка до современного человека, и столько же – до современного шимпанзе, то есть современники удалены друг от друга суммарно на 10 миллионов лет.
Используя эту константу скорости мутаций можно рассчитывать, сколько времени потребуется для прохождения определённого количества мутаций во фрагменте Y-хромосомы определённого размера. Например, мутация во фрагменте Y-хромосомы размером 8.47 миллионов нуклеотидов происходит в среднем раз в 144 года» [12, с. 38–40]. См. также: [13].
[2] Ср.: В. Н. Харьков: «За три последних десятилетия накоплено огромное количество данных, полученных при анализе нерекомбинирующей части Y-хромосомы. Первые работы в этой области относятся к середине 1980-х гг. В последующие годы маркеры Y-хромосомы стали использоваться для эволюционных исследований, в судебно-медицинской экспертизе, медицинской генетике и реконструкции родословных. Обнаружение диаллельных ДНК-маркеров на различных участках Y-хромосомы позволило открыть и классифицировать монофилетические гаплогруппы и начать подробное изучение генофонда различных популяций» [14, с. 982–983].
[3] Ср.: В. Н. Харьков: «Кроме характеристики генетического разнообразия, дифференциации и компонентного состава популяционного генофонда, исследование маркеров Y-хромосомы очень важно для филогенетических реконструкций. Использование множества различных полиморфных сайтов позволяет прослеживать комбинации аллелей, представляющие собой последовательную запись мутаций в ряду поколений. При этом под линией понимают группу гаплотипов, связанных общим происхождением, где каждый вариант отличается от соседнего на один мутационный шаг. Анализ филогении и филогеографии всего Y-хромосомного древа человечества, его отдельных клад и гаплогрупп является сутью этой области исследований.
Частота возникновения SNP на Y-хромосоме примерно в 2 раза выше, чем в среднем по геному, и уступает по скорости мутирования только мтДНК. Поскольку нерекомбинирующая часть Y-хромосомы значительно превосходит мтДНК по размеру, а число повторных и обратных мутаций для отдельных позиций намного ниже, то филогения мужских линий может быть гораздо более детально структурирована по сравнению с женскими. Накопление всё большего количества просеквенированных образцов Y-хромосом из различных популяционных выборок для всех гаплогрупп позволяет увеличивать информативность и популяционную специфичность их филогенетической реконструкции от самых древних линий до недавно возникших в локальных популяционных группах» [14, с. 983].
[4] Ср.: А. П. Деревянко: «Около 6–7 млн л.н. предковая линия человека в отряде приматов разделилась на две ветви – высших человекообразных обезьян и австралопитековых. В дальнейшем эволюционное развитие австралопитековых, которые расселялись только в Африке, происходило по сапиентной линии. Среди австралопитековых были группы ставших предковыми для рода Homo, первые представители сформировались ок. 2,8 млн л.н. Как показало исследование антроплогических находок, в позднем плиоцене – раннем плейстоцене на территории Африки существовали три вида рода Homo: H. rudolfensis, H. ergaster/erectus и H. habilis. Около 1,8 (1,7) млн л.н. H. ergaster/erectus вышли из Африки и стали расселяться в Евразии. Политипический вид H. erectus в ходе длительного и сложного эволюционного развития послужил основой для формирования людей современного типа – H. s. sapiens [Деревянко, 2012, 2017, 2019]. <…>
В результате эволюционного развития на предковой основе H. erectus в Африке 1,8–0,8 млн л.н. сформировался новый таксон, который получил у антропологов два названия – H. rhodensiensis и H. heidelbergensis. Морфологически и генетически популяции этих людей относились к одному биологическому виду, но их дальнейшая эволюционная история была разной. Homo rhodensiensis остались в Африке, на их предковой основе 200–150 тыс. л.н. сформировались люди современного типа (H. s. africaniensis). Homo heidelbergensis с ашельской индустрией ок. 800 тыс. л.н. мигрировали в Евразию (стоянка Гешер-Бенот-Яаков в Израиле). С этой миграцией связан первый (начальный) этап становления трёх таксонов – людей современного типа в Африке, неандертальцев и денисовцев в Евразии. Это подтверждается генетическими исследованиями: разделение общего предкового таксона на H. sapiens, с одной стороны, и H. s. neanderthalensis c H. s. altaiensis – с другой, произошло ок. 800 тыс. л.н. [Meyer et al., 2012]. Часть H. heidelbergensis с ашельской индустрией 700 (600) тыс. л.н. мигрировала в Европу, где в результате ассимиляционныз процессов с поздними эректусами (H. antecessor) через промежуточные формы Мауэр, Монморен, Штейнгейм, Араго 21, Сима-де-лос-Уэсос, Петралона и др. 200–150 тыс. л.н. сформировались классические неандертальцы с мустьерской индустрией. <…>
В начале верхнего плейстоцена, 120–60 тыс. л.н., в Африке и Евразии расселялись три таксона – люди современного типа в Африке (H. s. africaniensis), неандертальцы в Европе (H. s. neanderthalensis), денисовцы в Центральной и Северной Азии (H. s. altaiensis) [Деревянко 2012; и др.]. Представители этих таксонов скрещивались друг с другом, у них рождалось репродуктивное потомство. Это означает, что скрещивание происходило не между подвидами, а внутри одного вида. Если на финальном этапе эволюции рода Homo было три таксона с открытой генетической системой, то и на протяжении всей более чем 2,5-миллионнолетней эволюции у человека существовала открытая генетическая система, которая позволяла представителям таксонов скрещиваться, в результате чего рождалось репродуктивноспособное потомство. Все т.н. виды, которые были выделены антрополгами на основе небольшого количества останков со стоянок ранненго и среднего палеолита в Африке и Евразии, являлись подвидами с открытой генетической системой. Как показывают генетические исследования, у современных людей (не африканцев) в геноме сохраняется 1–2% генетического наследия неандертальцев. В геноме современных жителей Австралии и Океании имеется до 3–6% генетического наследия денисовцев [Reich et al., 2011]. Следовательно, неандертальцы и денисовцы при стволовой роли ранних людей современного анатомического типа, сформировавшихся в Африке 200–150 тыс. л.н., в процессе миграции последних в Евразию 80–50 тыс. л.н. внесли свой вклад в генетику и морфологию человека современного типа [Деревянко, 2012, 2019, 2022; Деревянко, Шуньков, Козликин, 2020].
В Восточной и Юго-Восточной Азии процесс сапиентного развития гоминин шёл с первончального заселения этих территорий H. erectus 1,7–1,6 млн л.н.; к настоящему времени здесь обнаружено ок. 10 антроплогических окаменелостей, относящихся к 120–60 тыс. л.н., которые антропологи связывают с людьми современного типа. Следует согласиться с мнением китайских исследователей о том, что в этих частях Азии сформировался четвёртый подвид человека современного типа (H. s. orientalensis), который также принял участие в формировании человека современного типа – H. s. sapiens [Деревянко, 2011]. <…>
…у денисовцев и неандертальцев был один предковый таксон – гейдельбергцы. В ходе миграции гейдельбергцев с ашельской индустрией в Европу 700 тыс. л.н. и ассимиляционных процессов с поздними эректусами (H. antecessor) при формировании классических неандертальцев (H. s. neanderthalensis) у последних сохранялась часть предкового генетического наследия. Доказательством этого являются выделенные у особи древностью ок. 430 тыс. лет из Сима-де-лос-Уэсос мтДНК денисовца и ядерная ДНК неандертальца [Meyer et al., 2014]. У гейдельбергцев, которые мигрировали на восток Азии значительно позже (400–350 тыс. л.н.) и ассимилировали поздних эректусов в Центральной Азии, что привело к формированию денисовцев (H. s. altaiensis), также сохранялась часть предкового генетического наследия, о чём свидетельствует мтДНК, выдлененная из культуросодержащего слоя 14 с денисовской индустрией. Это означает, что гейдельбергцы, расселившиеся на Ближнем Востоке, в Европе, Центральной Азии и на Алтае, были таксоном в процессе разделения на людей современного типа, неандертальцев и денисовцев и у них оставались открытая генетическая система, способность к скрещиванию, а также у тех и других – часть предкового генетического наследия» [16, с. 9, 11, 12, 13].
[5] Ср.: Р. Докинз: «Важная особенность ДНК заключается в том, что, пока цепь жизни не прервётся, закодированная в ДНК информация будет копироваться в новой молекуле ещё до разрушения старой. Поэтому информация живёт гораздо дольше молекул. Она возобновляется посредством копирования, а поскольку копирование для большинства “букв” является точным, теоретически она может сохраняться неопределённо долго. Значительная доля информации ДНК наших предков дошла до нас в неизменном виде, пережив в некоторых случаях сотни миллионов лет.
Таким образом, информация в ДНК – невероятно щедрый подарок, который природа преподнесла историкам. Какой историк мог надеяться, что каждая особь каждого вида носит в теле подробный документ! Более того, в этом тексте происходят незначительные случайные изменения, которые достаточно редки, чтобы не нарушить точность документа, но при этом достаточно часты, чтобы создать метки» [1, с. 33–34].
Ср. также: Н. Уэйд: «Как хранилище наследственной информации, пребывающей в постоянном изменении, геном подобен документу, который без конца переписывают. Но геном, изменяясь, сохраняет сведения обо всех «черновиках», которые содержат в себе хронику миллионов лет. Таким образом, геном может многое рассказать нам о разных временных пластах» [17, с. 8].
[6] Ср.: Э. Шрёдингер: «Важным, действительно судьбоносным событием в процессе размножения особи является не оплодотворение, а мейоз. Один набор хромосом достаётся от отца, другой – от матери. Ни случай, ни рок не могут появлиять на это» [18, с. 45].
[7] Ср.: В. П. Алексеев: «Весь антропогенез есть процесс накопления информации и уменьшения энтропии… Чем более высокие и развитые по своей организации формы создаёт эволюция, тем уже сфера энтропии и тем шире область информации» [19, с. 212].
Ср. также: Э. Шрёдингер: «Каждый процесс или событие – назовите, как хотите, – всё, что происходит в природе, означает повышение энтропии той части света, где это происходит. Так. Живой организм непрерывно повышает свою энтропию – или, можно сказать, проихводит положительную энтропию, – тем самым стремясь к опасному состоянию максимальной энтропии, то есть смерти. Чтобы держаться подальше от этого состояния – то есть жить, – организму необходимо постоянно черпать из окружающей среды отрицательную энтропию, которая, как мы сейчас поймём, а действительности является положительной. Организм питается отрицательной энтропией. Или, если выражаться яснее, важная особенность метаболизма заключается в том, что организму удаётся избавиться от всей той энтропии, какую он производит в ходе своей жизнедеятельности. <…> Теперь неуклюжее выражение «отрицательная энтропия» можно перефразировать более удачно: энтропия с отрицательным знаком есть мера порядка. Поэтому способ, которым организм постоянно поддерживает весьма высокий уровень упорядоченности (= весьма низкий уровень энтропии), в действительности заключается в непрерывном потреблении упорядоченности из окружающей среды. Этот вывод не столь парадоксален, сколь кажется на первый взгляд. Скорее его можно упрекнуть в тривиальности. На самом деле, в случае высших животных мы прекрасно знаем, какую упорядоченность они потребляют. Речь идёт о высокоупорядоченном состоянии вещества в относительно сложных органических соединениях, которые служат им пищей. После использования животные возвращают вещество в деградированном виде – однако не в полностью деградированном, поскольку растения могут употреблять его. Естественно, растения получают мощную дозу отрицательной энтропии в виде солнечного света» [18, с. 112, 115–116].
[8] Ср.: Д. Гриббин, М. Гриббин: «Процесс эволюции путём естественного отбора требует, чтобы живые существа воспроизводились, создавая себе подобных, но так, чтобы это копирование не было идеально точным, а порождало некоторое разнообразие в следующем поколении. Если среди этого разнообразия появляются отпрыски, которые по какой-то причине оказались успешнее остальных в деле выживания и воспроизводства, то признаки, обеспечившие их успешность, передаются следующим поколениям – иначе говоря, проходят отбор» [20, с. 7].
[9] Ср.: А. С. Пилипенко: «На самом деле митохондриальная Ева и Y-хромосомный Адам с точки зрения генетики даже не индивиды (люди), а лишь варианты их мтДНК и Y-хромосомы соответственно. Понятно, что носителями этих вариантов могла быть целая популяция того или иного размера» [23, с. 217]. Не согласимся с А. С. Пилипенко, который за пулом генов не видит конкретных индивидов – в частности, носителей SNPs. Несмотря на тот факт, что митохондриальная Ева и Y-хромосомный Адам находятся в постоянном дрейфе из-за накопления базы генетических данных, всякий раз речь идёт о конкретных индивидах, у которых эти мутации произошли, то есть о реальных предках, которых мы смогли реконструировать на основе текущих баз генетических данных, включая палеоДНК.
Ср.: Б. Сайкс: «В нас все перемешано – и в то же время мы все родственны друг другу. От каждого гена можно провести линию в прошлое, к другому общему предку. Это совершенно потрясающее, уникальное наследие, полученное нами от людей, живших раньше нас. Наши гены не возникают из ничего в момент нашего появления на свет – их несли нам, передавая через тысячи поколений, миллионы отдельных людей, конкретных личностей» [5, с. 301].
Ср.: А. А. Клёсов: «В связи с этим возникакает вопрос – а когда жил общий предок Homo sapiens, общий предок современных людей? Тот, который на самом верху обозначен как Y Root? Можно ли это время рассчитать? Ответ – можно, но с определённой степенью допущений. Первое и самое главное допущение – что общий предок сегодняшних людей на Земле на самом деле существовал. Естественно, речь не идёт о том, что на Земле когда-то жил один мужчина, и с ним рядом была одна женщина. Такого никогда не было. Во все времена было множество мужчин и множество женщин, а до них было множество самцов и самок, предшественников тех самых мужчин и женщин, и переход одних в других занял миллионы лет, и никто не может дать чётких определений, в чём выражался этот переход, это всё вопросы определений, которых, в общем-то, нет. Да, наверное, никого это отсутствие чётких определений особенно и не беспокоит.
Чисто концептуально, «общий предок» всех людей на сегодняшней планете – это тот, потомство которого выжило, в отличие от множества его современников, и продолжилось в потомках до настоящего времени. Помимо этого – это был тот человек, у которого было как минимум два сына, потомство которых выжило до настоящего времени. Как следствие первых двух условий, все потомки унаследовалиснип-мутации своего первопредка», и добавили снип-мутации в последующем. Поэтому есть ещё одно условие, точнее, следствие – мутации в гаплотипах потомково при экстраполяции в прошлое сходятся к «первопредку». Таким образом, можно определить его гаплотип, назависимо от того, как давно он жил» [12, с. 46–47].
Ср. также: «As with "Mitochondrial Eve", the title of "Y-chromosomal Adam" is not permanently fixed to a single individual, but can advance over the course of human history as paternal lineages become extinct. <…> Due to the definition via the "currently living" population, the identity of a MRCA, and by extension of the human Y-MRCA, is time-dependent (it depends on the moment in time intended by the term "currently"). The MRCA of a population may move forward in time as archaic lineages within the population go extinct: once a lineage has died out, it is irretrievably lost. This mechanism can thus only shift the title of Y-MRCA forward in time. Such an event could be due to the total extinction of several basal haplogroups. The same holds for the concepts of matrilineal and patrilineal MRCAs: it follows from the definition of Y-MRCA that he had at least two sons who both have unbroken lineages that have survived to the present day. If the lineages of all but one of those sons die out, then the title of Y-MRCA shifts forward from the remaining son through his patrilineal descendants, until the first descendant is reached who had at least two sons who both have living, patrilineal descendants. The title of Y-MRCA is not permanently fixed to a single individual, and the Y-MRCA for any given population would himself have been part of a population which had its own, more remote, Y-MRCA. <…> By the nature of the concept of most recent common ancestors, these estimates can only represent a terminus ante quem ("limit before which"), until the genome of the entire population has been examined (in this case, the genome of all living humans)» [21].
[10] Ср.: Р. Докинз: «Здесь я чувствую себя настолько неуверенно, что больше не буду пытаться оценивать число «пра-» перед «прародитель». Очень скоро счёт «пра-» пойдёт на миллиарды. Немного надёжнее поряжок ветвления линий, но и он может оказаться ошибочным» [1, с. 520].
[11] Ср.: «Исследование генома X-хромосомы в 2008 году привело к выводу, что азиатские популяции Homo erectus вполне могли скрещиваться с Homo sapiens и быть предками современных людей по смешанным линиям (не прямой мужской и не прямой женской)» [24].
[12] Ср.: А. А. Клёсов: «…общий предок современных гаплотипов обычно не тот, что был в «самом начале», а тот, прямые потомки которого выжили до настоящего времени. Потому и важно рассматривать данные ДНК-генеалогии в совокупности с данными археологии и лингвистики, причём именно данными, а не интерпретациями» [25, с. 30].
[13] Ср.: «Сравнение Y-хромосом двух денисовцев (Denisova4 (55–84 тыс. л. н.) и Denisova8 (106–136 тыс. л. н.)) с Y-хромосомами трёх неандертальцев и с Y-хромосомами современных неафриканских людей показало, что Y-хромосомная линия денисовцев отделились от Y-хромосомной линии современного человека около 700 тыс. л. н., Y-хромосомная линия неандертальцев отделилась от Y-хромосомной линии современного человека около 350 тыс. л. н. Для времени жизни общего предка носителей Y-хромосомной гаплогруппы А00 и носителей неафриканских Y-хромосомных гаплогрупп получили дату 249 тыс. лет назад» [28]. См.: [29].
[14] Ср.: «По данным сравнения Y-хромосомы неандертальца из пещеры Эль-Сидрон и африканца с Y-хромосомной гаплогруппой A00 время разделения линий неандертальцев и современных людей оценили по Y-хромосоме в 588 тыс. лет назад (95% доверительный интервал: 806–447 тыс. лет назад), а время появления Y-хромосомного Адама – в 275 тыс. лет назад (95% доверительный интервал: 304–245 тыс. лет назад)» [28] См.: [33],[34].
[15] Не исключено, что оба нуклеотида могут замениться по сравнению с предком – например, у денисовца и у неандертальца по сравнению с шимпанзе. Статистическая вероятность здесь – 1:62000000 (геном Y-хромосомы).
[16] Ср.: А. А. Клёсов: «За предковые (исходные) нуклеотиды принимали те, которые идентифицировали в Y-хромосомах мужчин планеты как неизменные (предковые), и подтверждённые по Y-хромосоме шимпанзе» [12, с. 46].
[17] См.: [36].
[18] Ср.: В. Н. Харьков: «Современная номенклатура Y-хромосомных линий представляет собой многоуровневую буквенно-цифровую систему обозначений, кластеризующихся в соответствии с пошаговой мутационной моделью. Иерархическое местоположение конкретной линии определяется сцеплением мутантного и исходного вариантов ДНК-маркера с лежащими выше и ниже SNP. Для обозначения субклад используются две системы: либо на основе буквенно-цифровой номенклатуры (например, E1b1, N1a1a1a1a2, R1a1a1b2a2, R1a1a1b2a2a3c, T1a2b и др.), либо по номеру терминальной мутации, определяющей данную гаплогруппу (например, Q-BZ99 или N-B172)» [14, с. 983].
[19] Ср.: В. Н. Харьков: «В момент возникновения гаплогруппы она представлена только у одного индивида, поскольку определяющая её мутация происходит на конкретной хромосоме, а затем частота гаплогруппы начинает стохастически меняться. Новая возникшая сублиния с большой вероятностью может исчезнуть из популяции, если мужской род, ведущий своё начало от родоначальника этой мутации, прервётся в каком-либо поколении. Большинство таких новых линий элиминируются стохастическим процессом генетического дрейфа. Но некоторые из гаплогрупп остаются в популяции и увеличивают свою частоту. На фоне стохастического роста численности гаплогруппы в ней с течением времени начинает накапливаться изменчивость по другим SNP и микросателлитным локусам. Изначально гаплогруппа представлена одним определённым гаплотипом-основателем, характеризующимся определённым значением аллелей в различных STR-локусах. Затем, с течением времени, у потомков появляются и другие аллели за счёт мутаций и возникают кластеры эволюционно родственных гаплотипов» [14, с. 984]. См. также: [37].
[20] Ср.: «The Y chromosome is passed from father to son remaining mostly unaltered across generations, except for small traceable changes in DNA. By tracking these changes, we constructed a family tree of humankind where all male lineages trace back to a single common ancestor who lived hundreds of thousands of years ago. This human tree allows us to explore lineages through time and place and to uncover the modern history of your direct paternal surname line and the ancient history of our shared ancestors» [30].
[21] Ср.: В. Н. Харьков: «Не менее значимым для классификации гаплогрупп является ресурс YFull.com, который был основан в 2013 г. и является сервисом по биоинформатическому анализу исходных данных секвенирования. Древо гаплогрупп на этом сайте является сейчас наиболее подробным и включает максимальное количество SNP, с привязкой к конкретным образцам. Обозначение линий при этом отличается от буквенно-цифровой кодировки и содержит только привязку к конкретным маркерам, начиная от самых базовых до терминальных для конкретного образца [10]» [14, с. 983].
[22] Ср.: «Subclade "formed" age: The TMRCA of a subclade is used as the "formed" age of each branch of the subclade. In other words, the formed age of a branch is the same as the TMRCA of the "parent" subclade of the branch» [46].
[23] Ср.: А. А. Клёсов: «ДНК-генеалогия выстраивает чёткую генеалогическую линию предков и потомков, как людей, принадлежащих к определённому роду. Поскольку гаплогруппа – это совокупность родственных субкладов, происходящих по цепочке от ещё более древних общих предков, а субклад – это совокупность носителей одних и тех же снипов, то ясно, что все их носители, то есть в данном случае мужчины, произошли от одного общего предка, патриарха, в ДНК которого этот снип впервые образовался. Фактически это есть общепринятое определение рода, что есть совокупность всех поколений людей, происходящих от одного предка.
Можно сказать и по-другому: гаплогруппа – совокупность гаплотипов, объединённая «групповой» необратимой мутацией, присущей определённому человеческому роду, то есть потомкам одного «патриарха», как правило, тысячелетия назад. Понятие «гаплогруппа» эквивалентно понятию «род» в ДНК-генеалогии. Эти мутации («снипы») выбирают по определённым критериям. Гаплогруппой также называют сам род в таких выражениях, как «гаплогруппа мигрировала шесть тысяч лет назад на восток», понимая, естественно, что мигрировали носители данной гаплогруппы. В настоящее время классификация включает 20 основных гаплогрупп (плюс А0 и А00), от А до Т в алфавитном порядке, и тысячи «нисходящих» гаплогрупп и субкладов.
Гаплогруппы не просто соответствуют своим родам, но образуют определённую последовательность, лестницу гаплогрупп, показывающих их иерархию – последовательный, ступенчатый переход от точки расхождения африканских и неафриканских популяций (примерно 160 тыс. лет назад) до самой недавней гаплогруппы R, образовавшейся примерно 30 тыс. лет назад. Эта лестница называется филогенетическим деревом гаплогрупп и их снипов. Все гаплогруппы и субклады на дереве должны включать снипы «вышестоящих» гаплогрупп и субкладов. То есть принцип «лесенки» должен выполняться. Преемственность узловых родов человечества должна соблюдаться» [12, с. 30–31].
[24] См.: [46]. См. также: [47].
[25] Ср.: «Ancestral Path. Every living man in the world shares a direct paternal line ancestor who lived around 230,000 years ago, and before that, our closest relatives are the Neanderthals and Denisovans. Each step represents a genetic ancestor on your direct paternal line. As more people test, this path will be further refined, and we will identify more steps on your ancestral line» [30].
[26] Референс Hg38 основан на представителе гаплогруппы R1b (R-S26186), поэтому все восходящие мутации, начиная с R1, сраниваются с шимпанзе (in the latest version of the human reference genome, GRCh38 released in 2017, the anonymous RP11 man accounts for most if not all of the Y-chromosome reference sequence [30]).
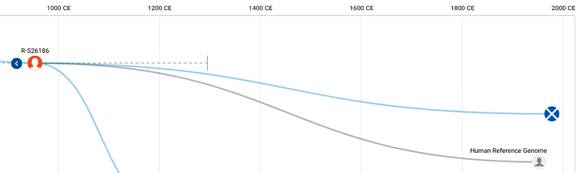
https://discover.familytreedna.com/y-dna/R-S26186/tree [30]
[27] См. также: [50],[51],[52],[53],[54],[55],[56],[57].
Библиография
1. Докинз Р. Рассказ предка. Паломничество к источкам жизни / Пер. с англ. С. Долотовской. М., 2021. 768 с.
2. Докинз Р. Перерастая Бога. Пособие для начинающих / Пер. с англ. А. Гопко. М., 2022. 256 с.
3. Докинз Р. Река, выходящая из Эдема. Жизнь с точки зрения дарвиниста / Пер. с англ. А. Гопко. М., 2020. 224 с.
4. Сайкс Б. Адамово проклятие. Мужчинам грозит медленное и неизбежное вымирание / Пер. с англ. Е. Я. Мигуновой. М., 2009.
5. Сайкс Б. Расшифрованный код Ледового человека: От кого мы произошли, или Семь дочерей Евы / Пер. с англ. Е. Я. Мигуновой. М., 2005. 304 с.
6. Винарский М. В. Евангелие от LUCA. В поисках родословной животного мира. М., 2021. 352 с.
7. Пэабо С. Неандерталец. В поисках исчезнувших геномов / Пер. с англ. Е. Наймарк. М., 2018. 416 с.
8. Шубин Н. Внутренняя рыба. История человеческого тела с древнейших времён до наших дней / Пер. с англ. П. Петрова. М., 2021. 304 с.
9. Кукушкин Н. В. Хлопок одной ладонью. Как неживая природа породила человеческий разум. М., 2021. 542 с.
10. Ястребов С. А. От атомов к древу. Введение в современную науку о жизни. М., 2018. 704 с.
11. Дарвин Ч. Р. Происхождение видов путём естественного отбора, или Сохранение благоприятных рас в борьбе за жизнь / Пер. с англ. К. А. Тимирязева, М. А. Мензбира, А. П. Павлова, И. А. Петровского. М., 2020. 592 с.
12. Клёсов А. А. ДНК-генеалогия от А до Т. М., 2016. 480 с.
13. What is YFull's age estimation methodology? / YFull: FAQ. [Эл. ресурс]. URL: https://www.yfull.com/faq/what-yfulls-age-estimation-methodology/ (Дата обращ. 01.07.2023).
14. Харьков В. Н. Маркеры Y-хромосомы в популяционной генетике: фундаментальные и прикладные результаты этногеномных исследований // Генетика. 2021. Т. 57. № 9. С. 981–994. URL: https://elibrary.ru/item.asp?id=46350988 (Дата обращ. 01.07.2023).
15. Homo sapiens isolate NA24385 chromosome Y / GenBank. [Эл. ресурс]. URL: https://www.ncbi.nlm.nih.gov/nuccore/CP086569.2 (Дата обращ. 01.07.2023).
16. Деревянко А. П. Исследование палеолита Сибири // Археология, этнография и антропология Евразии. 2023. Т. 51. № 1. С. 3–17.
17. Уэйд Н. На заре человечества. Неизвестная история наших предков / Пер. с англ. Н. Мезин. М., 2022. 408 с.
18. Шрёдингер Э. Что такое жизнь? / Пер. с англ. К. С. Егоровой. М., 2019. 288 с.
19. Пряхин В. Ф. Бессмертие – мировоззрение XXI века. М., 2018. 268 с.
20. Гриббин Д., Гриббин М. Происхождение эволюции: идея естественного отбора до и после Дарвина. М., 2022.
21. Y-chromosomal Adam / Wikipedia. [Эл. ресурс]. URL: https://en.wikipedia.org/wiki/Y-chromosomal_Adam (Дата обращ. 01.07.2023).
22. Ближайший общий предок / Википедия. [Эл. ресурс]. URL: https://ru.wikipedia.org/wiki/Ближайший_общий_предок (Дата обращ. 01.07.2023).
23. Естественнонаучные предметы. Практическая молекулярная генетика для начинающих: 8–9 классы / Уч. пос. под ред. П. М. Бородина и Е. Н. Ворониной. 2-е изд., стер. М., 2022. 272 с.
24. Человек прямоходящий / Википедия. [Эл. ресурс]. URL: https://ru.wikipedia.org/wiki/Человек_прямоходящий (Дата обращ. 01.07.2023).
25. Клёсов А. А. Народы России. ДНК-генеалогия. СПб., 2021. 784 с.
26. GeneticHomeland.com / Brad Larkin. [Эл. ресурс]. URL: https://www.genetichomeland.com, https://www.genetichomeland.com/welcome/dnapedigree.asp (Дата обращ. 01.07.2023).
27. International Society of Genetic Genealogy (ISOGG). [Эл. ресурс]. URL: https://isogg.org/tree/, https://isogg.org/tree/ISOGG_YDNATreeTrunk.html (Дата обращ. 01.07.2023).
28. Y-хромосомный Адам / Википедия. [Эл. ресурс]. URL: https://ru.wikipedia.org/wiki/ Y-хромосомный_Адам (Дата обращ. 01.07.2023).
29. Petr M., Hajdinjak M., Fu Q., Essel E., Rougier H., Crevecoeur I., Semal P., Golovanova L. V., Doronichev V. B., Lalueza-Fox C., de la Rasilla M., Rosas A., Shunkov M. V., Kozlikin M. B., Derevianko A. P., Vernot B., Meyer M., Kelso J.. The evolutionary history of Neandertal and Denisovan Y chromosomes // Science 369 (6511). 25 Sep 2020. P. 1653–1656. URL: https://www.science.org/doi/10.1126/science.abb6460 (Дата обращ. 01.07.2023).
30. DiscoverTM More (Beta) [Эл. ресурс]. URL: https://discover.familytreedna.com/, https://discover.familytreedna.com/y-dna/R-S26186/notable (Дата обращ. 01.07.2023).
31. Skov L., Peyrégne S., Popli D., Iasi L. N. M., Devièse T., Slon V., Zavala E. I., Hajdinjak M., Sümer A. P., Grote S., Mesa A. B., Herráez D. L., Nickel B., Nagel S., Richter J., Essel E., Gansauge M., Schmidt A., Korlević P., Comeskey D., Derevianko A. P., Kharevich A., Markin S. V., Talamo S., Douka K., Krajcarz M. T., Roberts R. G., Higham T., Viola B., Krivoshapkin A. I., Kolobova K. A., Kelso J., Meyer M., Pääbo S., Peter B. M. Genetic insights into the social organization of Neanderthals // Nature. V. 610. 20 October 2022. P. 519–525. URL: https://www.nature.com/articles/s41586-022-05283-y (Дата обращ. 01.07.2023).
32. YFull.com. [Эл. ресурс]. URL: https://www.yfull.com/ (Дата обращ. 01.07.2023).
33. Mendez F. L., Poznik G. D., Castellano S., Bustamante C. D. The Divergence of Neandertal and Modern Human Y Chromosomes // The American Journal of Human Genetics. 07 April 2016. V. 98. I. 4. P. 728–734. URL: https://www.cell.com/ajhg/fulltext/S0002-9297(16)30033-7 (Дата обращ. 01.07.2023).
34. Lowery R. K., Uribe G., Jimenez E. B., Weiss M. A., Herrera K. J., Regueiro M., Herrera R. J. Neanderthal and Denisova genetic affinities with contemporary humans: Introgression versus common ancestral polymorphisms // Gene. 1 November 2013. V. 530. I. 1. P. 83–94. URL: https://www.sciencedirect.com/science/article/abs/pii/S0378111913007567?via%3Dihub (Дата обращ. 01.07.2023).
35. Боринская С. А. Смешение неандертальцев, денисовцев и сапиенсов: спор разгорается с новой силой. [Эл. ресурс]. URL: http://antropogenez.ru/interview/709/ (Дата обращ. 01.07.2023).
36. Нилогов А. С. Снипование как именование (антиязыковая методология в помощь ДНК-генеалогии) // Litera. 2016. № 1. С. 18–25. [Эл. ресурс]. URL: http://e-notabene.ru/fil/article_17670.html (Дата обращ. 01.07.2023).
37. Харьков В. Н., Зарубин А. А., Вагайцева К. В., Раджабов М. О., Новикова Л. М., Валихова Л. В., Хитринская И. Ю., Степанов В. А. Y-хромосома как инструмент для ДНК-идентификации и определения популяционный принадлежности // Генетика. 2020. Т. 56. № 9. С. 1065–1074. URL: https://www.elibrary.ru/item.asp?id=43181768 (Дата обращ. 01.07.2023).
38. Family Tree DNA. [Эл. ресурс]. URL: https://www.familytreedna.com (Дата обращ. 01.07.2023).
39. Nebula Genomics. [Эл. ресурс]. URL: https://nebula.org/whole-genome-sequencing-dna-test (Дата обращ. 01.07.2023).
40. YFull YTree. [Эл. ресурс]. URL: https://www.yfull.com/tree, https://www.yfull.com/check-snp (Дата обращ. 01.07.2023).
41. ISOGG YBrowse / Human Y Chromosome Browser (based on hg38). [Эл. ресурс]. URL: http://ybrowse.org, https://ybrowse.org/gb2/gbrowse_details/chrY?ref=chrY;start=7684315;end=7684315;name=Z93;class=Sequence;feature_id=50189;db_id=chrY%3Adatabase (Дата обращ. 01.07.2023).
42. Mendez F. L., Krahn T., Schrack B., Krahn A.-M., Veeramah K. R., Woerner A. E., Fomine F. L. M., Bradman N., Thomas M. G., Karafet T. M., Hammer M. F. An African American paternal lineage adds an extremely ancient root to the human Y chromosome phylogenetic tree // The American Journal of Human Genetics. 07 March 2013. V. 92. I. 3. P. 454–459. URL: https://www.cell.com/ajhg/fulltext/S0002-9297(13)00073-6 (Дата обращ. 01.07.2023).
43. YTree Dnachron. [Эл. ресурс]. URL: https://www.dnachron.com/ytree (Дата обращ. 01.07.2023).
44. YTree Mofang. [Эл. ресурс]. URL: https://www.23mofang.com/ancestry/ytree/root (Дата обращ. 01.07.2023).
45. Y-Haplotree FTDNA. [Эл. ресурс]. URL: https//www.familytreedna.com/public/y-dna-haplotree/A (Дата обращ. 01.07.2023).
46. What is YFull's subclade age methodology? / YFull: FAQ. [Эл. ресурс]. URL: https://www.yfull.com/faq/how-does-yfull-determine-formed-age-tmrca-and-ci/ (Дата обращ. 01.07.2023).
47. Adamov D., Guryanov V., Karzhavin S., Tagankin V., Urasin V. Defining a New Rate Constant for Y-Chromosome SNPs based on Full Sequencing Data // The Russian Journal of Genetic Genealogy. 2015. Т. 7. № 1. С. 68–89. URL: https://www.researchgate.net/publication/273773255_Defining_a_New_Rate_Constant_for_Y-Chromosome_SNPs_based_on_Full_Sequencing_Data (Дата обращ. 01.07.2023).
48. Нилогов А. С. Генетико-генеалогическое (генетеалогическое) исследование рода Нилоговых (на материале ДНК аутосом и Y-хромосомы) // Учёные записки Минусинского краеведческого музея им. Н. М. Мартьянова. Вып. № 3. Минусинск, 2021. С. 161–212. URL: https://музей-мартьянова.рф/wp-content/uploads/2022/06/Учёные-записки-2.pdf (Дата обращ. 01.07.2023).
49. Нилогов А. С. Генетико-генеалогическое (генетеалогическое) исследование рода Нилоговых (на материале ДНК аутосом) // Genesis: исторические исследования. 2023. № 2. С. 113–131. [Эл. ресурс]. URL: http://e-notabene.ru/hr/article_37301.html (Дата обращ. 01.07.2023).
50. Мартынов М. Е., Нилогов А. С. Косинско-Камское поречье в переписях и ревизиях XVI – XIX вв. Кудымкар, 2012. 252 с.
51. Нилогов А. С. Откуда есть пошли Нилоговы // Генеалогический вестник. 2012. № 44. С. 27–55.
52. Нилогов А. С. ДНК-генеалогия на службе у генеалогии // The Russian Journal of Genetic Genealogy (русская версия). 2015. Т. 7. № 2. С. 51–66.
53. Нилогов А. С. ДНК-генеалогия на службе у генеалогии // Ада чир-суу = Отечество: краеведческий альманах. Сост. М. А. Аева. Вып. 3. Абакан, 2015. С. 78–92. URL: https://nbdrx.ru/pdf/almanac/Ada_Chir-Suu_V-003-2015.pdf (Дата обращ. 01.07.2023).
54. Нилогов А. С. Генетика как дополнительный инструмент генеалогии // Генеалогический вестник. 2017. № 54. С. 85–91. URL: https://geno.ru/media/files/генеалогический_вестник/гв-54.pdf, https://geno.ru/media/files/генеалогический_вестник/гв-54_статистический_материал_к_статье_а.с._нилогова.pdf (Дата обращ. 01.07.2023).
55. Нилогов А. С. Семь колен восходящей родословной Алексея Сергеевича Нилогова. Германия: Саарбрюккен, 2017. 388 с. URL: https://yadi.sk/d/xXh9q9fN3Qzczh (Дата обращ. 01.07.2023).
56. Нилогов А. С. До седьмого колена… Германия: Саарбрюккен, 2019. 356 с. URL: https://yadi.sk/i/rDcUisWcoVQwjw (Дата обращ. 01.07.2023).
57. Нилогов А. С. До восьмого колена… Германия: Саарбрюккен, 2020. 438 с. URL: https://yadi.sk/d/bB8Pppu8XbdFPw (Дата обращ. 01.07.2023).
58. Big Y Block Tree / FTDNA. [Эл. ресурс]. URL: https://www.familytreedna.com/my/bigy-blocktree (Дата обращ. 01.07.2023).
59. Нилогов А.С. Секвенирован первый полный геном жителя Хакасии / Абакан. № 34. 01–07 сентября 2021. С. 19. URL: https://nbdrx.ru/pdf/Abakan/2021/gab2021034.pdf (Дата обращ. 01.07.2023).
60. Известный учёный из Хакасии расшифровал свой геном в американской лаборатории / Новая Хакасия. 20.12.2021. [Эл. ресурс]. URL: https://newkhakasiya.online/news/2021-12-20/izvestnyy-uchenyy-iz-hakasii-rasshifroval-svoy-genom-v-amerikanskoy-laboratorii-1083322 (Дата обращ. 01.07.2023)
References
1. Dawkins, R. (2021). The Ancestor's Story. Pilgrimage to the sources of life. Moscow: AST, CORPUS.
2. Dawkins, R. (2022). Outgrowing God. Handbook for beginners. Moscow: AST, CORPUS.
3. Dawkins, R. (2020). The river coming out of Eden. Life from the point of view of a Darwinist. Moscow: AST, CORPUS.
4. Sykes, B. (2009). Adam's curse. Men are threatened with slow and inevitable extinction. Moscow: RIPOL classic.
5. Sykes, B. (2005). The deciphered code of the Ice Man: From whom we came, or the Seven Daughters of Eve. Moscow: RIPOL classic.
6. Vinarsky, M. V. (2021). The Gospel of LUCA. In search of the pedigree of the animal world. Moscow: Alpina non-fiction.
7. Peabo, S. (2018). Neanderthal. In search of disappeared genomes. Moscow: AST, CORPUS.
8. Shubin, N. (2021). Internal fish. The history of the human body from ancient times to the present day. Moscow: AST, CORPUS..
9. Kukushkin, N. V. (2021). Clap with one palm. How inanimate nature gave birth to the human mind. Moscow: Alpina non-fiction.
10. Yastrebov, S. A. (2018). From atoms to the tree. Introduction to the modern science of life. Moscow: Alpina non-fiction.
11. Darwin, C. R. (2020). The origin of species by natural selection, or the preservation of favorable races in the struggle for life. Moscow: AST.
12. Klesov, A. A. (2016). DNA genealogy from A to T. Moscow: Book word.
13. What is YFull's age estimation methodology? YFull: FAQ. [email resource]. Retrieved from https://www.yfull.com/faq/what-yfulls-age-estimation-methodology
14. Kharkov, V. N. (2021). Markers of the Y chromosome in population genetics: fundamental and applied results of ethnogenomic studies. Genetics, 57: 9, 981–994. doi:10.31857/S0016675821090046 Retrieved from https://elibrary.ru/item.asp?id=46350988
15. Homo sapiens isolate NA24385 chromosome Y. GenBank. [Electronic resource]. Retrieved from https://www.ncbi.nlm.nih.gov/nuccore/CP086569.2
16. Derevyanko, A. P. (2023). Investigation of the Paleolithic of Siberia. Archeology, ethnography and anthropology of Eurasia, 51:1, 3–17. doi:10.17746/1563-0102.2023.51.1.003-017
17. Wade, N. (2022). At the dawn of humanity. The unknown history of our ancestors. Moscow: Alpina non-fiction.
18. Schrodinger, E. (2019). What is life? Moscow: AST.
19. Pryakhin, V. F. (2018). Immortality – the worldview of the XXI century.Moscow: Academic project.
20. Gribbin D., Gribbin M. (2022). The origin of evolution: the idea of natural selection before and after Darwin. Moscow: Alpina non-fiction.
21. Y-chromosomal Adam / Wikipedia. [Electronic resource]. Retrieved from https://en.wikipedia.org/wiki/Y-chromosomal_Adam
22. The closest common ancestor / Wikipedia. [Electronic resource]. Retrieved from https://ru.wikipedia.org/wiki/Ближайший_общий_предок
23. Natural science subjects. Practical molecular genetics for beginners: grades 8-9. (2022). Moscow: Education.
24. The man is upright / Wikipedia. [Electronic resource]. Retrieved from https://ru.wikipedia.org/wiki/Человек_прямоходящий
25. Klesov, A. A. (2021). Peoples of Russia. DNA genealogy. St. Petersburg: а.
26. GeneticHomeland.com Brad Larkin. [Electronic resource]. Retrieved from https://www.genetichomeland.com , https://www.genetichomeland.com/welcome/dnapedigree.asp
27. International Society of Genetic Genealogy (ISOGG). [Electronic resource]. Retrieved from https://isogg.org/tree /, https://isogg.org/tree/ISOGG_YDNATreeTrunk.html
28. Y-chromosomal Adam / Wikipedia. [Electronic resource]. Retrieved from https://ru.wikipedia.org/wiki / Y-chromosomal_Adam
29. Petr, M., Hajdinjak, M., Fu, Q., Essel, E., Rougier, H., Crevecoeur, I., Semal, P., Golovanova, L. V., Doronichev, V. B., Lalueza-Fox, C., de la Rasilla, M., Rosas, A., Shunkov, M. V., Kozlikin, M. B., Derevianko, A. P., Vernot, B., Meyer, M., & Kelso, J. (2020). The evolutionary history of Neandertal and Denisovan Y chromosomes. Science, 369, 1653-1656. doi:10.1126/science.abb6460 Retrieved from https://www.science.org/doi/10.1126/science.abb6460
30. DiscoverTM More (Beta) [Electronic resource]. Retrieved from https://discover.familytreedna.com /, https://discover.familytreedna.com/y-dna/R-S26186/notable
31. Skov, L., Peyrégne, S., Popli, D., Iasi, L. N. M., Devièse, T., Slon, V., Zavala, E. I., Hajdinjak, M., Sümer, A. P., Grote, S., Mesa, A. B., Herráez, D. L., Nickel, B., Nagel, S., Richter, J., Essel, E., Gansauge, M., Schmidt, A., Korlević, P., Comeskey, D., Derevianko, A. P., Kharevich, A., Markin, S. V., Talamo, S., Douka, K., Krajcarz, M. T., Roberts, R. G., Higham, T., Viola, B., Krivoshapkin, A. I., Kolobova, K. A., Kelso, J., Meyer, M., Pääbo, S., & Peter, B. M. (2022). Genetic insights into the social organization of Neanderthals. Nature, 610, 519–525. doi:https://doi.org/10.1038/s41586-022-05283-y Retrieved from https://www.nature.com/articles/s41586-022-05283-y
32. YFull.com. [Electronic resource]. Retrieved from https://www.yfull.com
33. Mendez, F. L., Poznik, G. D., Castellano, S., & Bustamante, C. D. (2016). The Divergence of Neandertal and Modern Human Y Chromosomes. The American Journal of Human Genetics, 98:4, 728–734. doi:https://doi.org/10.1016/j.ajhg.2016.02.023 Retrieved from https://www.cell.com/ajhg/fulltext/S0002-9297(16)30033-7
34. Lowery, R. K., Uribe, G., Jimenez, E. B., Weiss, M. A., Herrera, K. J., Regueiro, M., & Herrera, R. J. (2013). Neanderthal and Denisova genetic affinities with contemporary humans: Introgression versus common ancestral polymorphisms. Gene, 530: 1, 83–94. doi:https://doi.org/10.1016/j.gene.2013.06.005 Retrieved from https://www.sciencedirect.com/science/article/abs/pii/S0378111913007567?via%3Dihub
35. Borinskaya, S. A. Mixing of Neanderthals, Denisovans and Sapiens: the dispute flares up with renewed vigor. [Electronic resource]. Retrieved from http://antropogenez.ru/interview/709
36. Nilogov, A. S. (2016). Sniping as naming (anti-linguistic methodology to help DNA genealogy). Litera, 1, 18-25. doi:10.7256/2409-8698.2016.1.17670 [Electronic resource]. Retrieved from http://e-notabene.ru/fil/article_17670.html
37. Kharkov, V. N., Zarubin, A. A., Vagaytseva, K. V., Radjabov, M. O., Novikova, L. M., Valikhova, L. V., Khitrinskaya, I. Yu., & Stepanov, V. A. (2020). The Y-chromosome as a tool for DNA identification and determination of population affiliation. Genetics, 56: 9, 1065–1074. doi:10.31857/S0016675820090118 Retrieved from https://www.elibrary.ru/item.asp?id=43181768
38. Family Tree DNA. [Electronic resource]. Retrieved from https://www.familytreedna.com
39. Nebula Genomics. [Electronic resource]. Retrieved from https://nebula.org/whole-genome-sequencing-dna-test
40. YFull YTree. [Electronic resource]. Retrieved from https://www.yfull.com/tree , https://www.yfull.com/check-snp
41. ISOGG YBrowse / Human Y Chromosome Browser (based on hg38). [Electronic resource]. Retrieved from http://ybrowse.org , https://ybrowse.org/gb2/gbrowse_details/chrY?ref=chrY;start=7684315;end=7684315;name=Z93;class=Sequence;feature_id=50189;db_id=chrY% 3Adatabase
42. Mendez, F. L., Krahn, T., Schrack, B., Krahn, A.-M., Veeramah, K. R., Woerner, A. E., Fomine, F. L. M., Bradman, N., Thomas, M. G., Karafet, T. M., & Hammer, M. F. (2013). An African American paternal lineage adds an extremely ancient root to the human Y chromosome phylogenetic tree. The American Journal of Human Genetics, 92: 3, 454–459. doi:https://doi.org/10.1016/j.ajhg.2013.02.002 Retrieved from https://www.cell.com/ajhg/fulltext/S0002-9297(13)00073-6
43. YTree Dnachron. [Electronic resource]. Retrieved from https://www.dnachron.com/ytree
44. YTree Mofang. [Electronic resource]. Retrieved from https://www.23mofang.com/ancestry/ytree/root
45. Y-Haplotree FTDNA. [Electronic resource]. Retrieved from https//www.familytreedna.com/public/y-dna-haplotree/A
46. What is YFull's subclade age methodology? / YFull: FAQ. [email resource]. Retrieved from https://www.yfull.com/faq/how-does-yfull-determine-formed-age-tmrca-and-ci
47. Adamov, D., Guryanov, V., Karzhavin, S., Tagankin, V., & Urasin, V. (2015). Defining a New Rate Constant for Y-Chromosome SNPs based on Full Sequencing Data. The Russian Journal of Genetic Genealogy, 7: 1, 68–89. Retrieved from https://www.researchgate.net/publication/273773255_Defining_a_New_Rate_Constant_for_Y-Chromosome_SNPs_based_on_Full_Sequencing_Data
48. Nilogov, A. S. (2021). Genetic and genealogical (genealogical) study of the genus Nilogov (based on the DNA of autosomes and Y-chromosomes). Scientific notes of the Minusinsk Museum of Local Lore named after N. M. Martyanov, 3, 161–212. Retrieved from https://музей-мартьянова .rf/wp-content/uploads/2022/06/ Scientists-notes-2.pdf
49. Nilogov, A. S. (2023). Genetic and genealogical (genealogical) study of the genus Nilogov (based on DNA autosomes). Genesis: historical research, 2, 113–131. doi: 10.25136/2409-868X.2023.2.37301. [Electronic resource]. Retrieved from http://e-notabene.ru/hr/article_37301.html
50. Martynov, M. E., & Nilogov, A. S. (2012). Kosinsko-Kama river in censuses and revisions of the XVI – XIX centuries. Kudymkar: Ethnocultural center.
51. Nilogov, A. S. (2012). Where did the Nilogovs come from. Genealogical Bulletin, 44, 27–55.
52. Nilogov, A. S. (2015). DNA genealogy in the service of genealogy. The Russian Journal of Genetic Genealogy (Russian version), 7:2, 51–66.
53. Nilogov, A. S. (2015). DNA genealogy in the service of genealogy. Ada chir-suu = Fatherland: local history almanac, 3, 78–92. Retrieved from https://nbdrx.ru/pdf/almanac/Ada_Chir-Suu_V-003-2015.pdf
54. Nilogov, A. S. (2017). Genetics as an additional tool of genealogy. Genealogical Bulletin, 54, 85–91. Retrieved from https://geno.ru/media/files/генеалогический_вестник/гв-54.pdf , https://geno.ru/media/files/генеалогический_вестник/гв-54_статистический_материал_к_статье_а .S._nilogova.pdf
55. Nilogov, A. S. (2017). Seven tribes of the ascending pedigree of Alexey Sergeevich Nilogov. Germany, Saarbrücken: Lambert Academic Publishing. Retrieved from https://yadi.sk/d/xXh9q9fN3Qzczh
56. Nilogov, A. S. (2019). To the seventh generation... Germany, Saarbrücken: Lambert Academic Publishing. Retrieved from https://yadi.sk/i/rDcUisWcoVQwjw
57. Nilogov, A. S. (2020). To the eighth generation... Germany, Saarbrücken: Lambert Academic Publishing. Retrieved from https://yadi.sk/d/bB8Pppu8XbdFPw
58. Big Y Block Tree / FTDNA. [Electronic resource]. Retrieved from https://www.familytreedna.com/my/bigy-blocktree
59. Nilogov, A. S. (2021). Sequenced the first complete genome of a resident of Khakassia. Abakan, 34, 19. Retrieved from https://nbdrx.ru/pdf/Abakan/2021/gab2021034.pdf
60. A famous scientist from Khakassia decoded his genome in an American laboratory. New Khakassia. 20.12.2021. [Electronic resource]. Retrieved from https://newkhakasiya.online/news/2021-12-20/izvestnyy-uchenyy-iz-hakasii-rasshifroval-svoy-genom-v-amerikanskoy-laboratorii-1083322
Результаты процедуры рецензирования статьи
В связи с политикой двойного слепого рецензирования личность рецензента не раскрывается.
Со списком рецензентов издательства можно ознакомиться здесь.
Отзыв
На статью « Y-хромосомные предки: к проблеме генетико-генеалогической номинации».
Предмет обозначен в названии и разъяснен в тексте статьи. В статье автор ставит цель изучить родословную Ниловых и степень родства его представителей (мужской и женской линии).
Методология исследования. Данная работа базируетс на комплексной генетико-генеалогической методологии.
Актуальность исследования. В последние десятилетия возрос и отмечает автор рецензируемой статьи «новый уровень ДНК-генеалогической (генетико-генеалогической, генетеалогической) верификации поставил под методологический вопрос классическое документально-архивное родословие, так как под родословной понимается прежде всего биологическое (генетическое, филогенетическое) родство» и написать данную статью автора подвигла книга британского дарвиниста Р.Докинза «Рассказ предка».
Новизна статьи заключается в постановке проблемы и задач исследования. Новизна заключается также в том, что в статье делается попытка изучить влияние Y-хромосомы в генетико-геналогической номинации. Научная новизна также заключается в том, что автор привлекает и архивные материалы по генеалогии рода Ниловых. Научная новизна определяется также привлечением архивных материалов.
Автор в своем исследовании использует « несколько генетических баз данных (Y-DNA database), среди которых наиболее авторитетными являются: YFull YTree , Discover Family Tree DNA, YSNP YTree от ISOGG « и др. Стиль статьи научный и для того, ясный и четкий. Автор в начале статьи дает разъяснение применяемых в статье терминов, что облегчает восприятие статьи гуманитариями и другими категориями читателей, кто может плохо разбираться в биологических терминах. В самом начале статьи автор представил цитаты из работ Р. Докинза ( 4 цитатыт), Б. сайкса ( 5 цитат), Н. Шубина ( 3 цитаты), по две цитаты Н.В. Кукушкина и С.А. Ястребова , по одной цитате С. Пэабо, Ч. Дарвина. Приведение данных цитат в статье помогает понять цели и задачи автора, а также содержание статьи в целом. После приведения данных цитат идет текст самой статьи. Текст логически выстроен, снабжен рисунками, графиками из других статей и электронных ресурсов.
Выводы автора логично вытекают из проведенной работы и автор констатирует, что «на примере генетико-генеалогической (генетеалогической) реконструкции патрилинии автора были показаны перспективы комплексного (междисциплинарного) исследования человеческого патрилинейного родства (см. также: ). Проблема номинации наших далёких предков рассмотрена в аспекте лингвофилософской проблемы ономафикации как части патрофикации – общего дела по воскрешению отцов (Н. Ф. Фёдоров). Лингвогенетеалогический метод снипования как именования (антропонимофикации) от Y-хромосомного Адама через все узловые ветви (гаплогруппы/снипы) к документально подтверждённой фактологии классической генеалогии должен стать решением насущной проблемы по реконструкции популяционной и персональной генеалогии людей». И в подтверждении своих выводов автор приводит ссылки на данные таких исследователей как А. А. Клёсов, В. Н. Харьков, А. П. Деревянко, Р. Докинз, Н. Уэйд, Э. Шрёдингер, Д. Гриббин, М. Гриббин, А. С. Пилипенко, Б. Сайкс и др.
Библиография статьи насчитывает 60 работ, в том числе и 11 работ автора статьи и одну статью о его работах. Библиография обширная, разноплановая и свидетельствует, как и статья в целом о том, что автор статьи глубоко интересуется темой, статья его несомненно оригинальная и интересная, вместе с тем, следует отметить, что она может вызвать дискуссию среди специалистов по теме в целом и отдельным положениям автора статьи.
Апелляция к оппонентам представлена на уровне собранной информации, полученной автором в ходе работы над темой статьи, другим работам автора, представленным в библиографии и в библиографии в целом.
Статья интересная, актуальная и, по мнению рецензента, она может вызвать дискуссию среди специалистов и неспециалистов, т.к. несмотря на то, что автор статьи постарался раскрыть терминологию статьи и статья написана ясным языком, но может вызвать сложности для восприятия. Статья может быть опубликована в журнале, она заслуживает внимания и вызовет интерес.
|







