|
Сельское хозяйство
Правильная ссылка на статью:
Рыжова Л.В. Онтогенез и динамика онтогенетической структуры ценопопуляций щирицы запрокинутой (Amaranthus retroflexus L.) // Сельское хозяйство. 2021. № 3. С. 22-30. DOI: 10.7256/2453-8809.2021.3.37212 URL: https://nbpublish.com/library_read_article.php?id=37212
Онтогенез и динамика онтогенетической структуры ценопопуляций щирицы запрокинутой (Amaranthus retroflexus L.)
Рыжова Людмила Валерьяновна
ORCID: 0000-0003-2512-7078
кандидат биологических наук
доцент кафедры биологии ФГБОУ ВО "Марийский государственный университет"
424000, Россия, республика Марий Эл, г. Йошкар-Ола, пл. Ленина, 1
Ryzhova Lyudmila
PhD in Biology
Associate Professor at the Department of Biology of Mari State University
424000, Russia, respublika Marii El, g. Ioshkar-Ola, pl. Lenina, 1

|
procopjeva@mail.ru
|
|
 |
|
DOI: 10.7256/2453-8809.2021.3.37212
Дата направления статьи в редакцию:
24-12-2021
Дата публикации:
31-12-2021
Аннотация:
Объектом исследования является щирица (амарант) запрокинутая (Amaránthus retrofléxus L.) – североамериканский вид, инвазионный вид для России, однолетнее сорное растение. Цель работы – изучение онтогенеза и онтогенетической структуры ценопопуляций щирицы запрокинутой. Исследования проводились в 2014 г. в окрестностях деревни Сизнерь Мари-Турекского района Республики Марий Эл на картофельном и свекольном полях. В пределах каждого поля было заложено по 10-15 учетных площадок размером 1м2, на которых велись еженедельные наблюдения за развитием растений щирицы. Для оценки плотности и онтогенетической структуры популяций проводили анализ таблиц сопряженности R×C с помощью точного критерия Фишера и критерия χ2 (Глотов, 1984). К основным выводам проведенного исследования можно отнести следующее. Впервые выделено пять онтогенетических состояний щирицы запрокинутой (от проростов до генеративных). Маркерами онтогенетических состояний являются качественные и количественные признаки: структура и ветвление побега, размеры и форма листовых пластинок, число боковых жилок листа длина корня, наличие боковых корней разных порядков. Плотность растений щирицы на обоих полях не различается и составляет в среднем 6,9 экз./м2. Изменение онтогенетической структуры ценопопуляций щирицы запрокинутой на исследуемых полях происходит по-разному. Анализ показал, что в пределах поля распределения по онтогенетическим состояниям между учетными площадками в первый и последний сроки наблюдения однородные (Р>0,13). В остальные сроки распределения по онтогенетическим состояниям на разных учетных площадках в пределах поля различные (Р˂0,00001). На свекольном поле растения проходят онтогенез быстрее. В середине июля на свекольном поле представлены все онтогенетические состояния щирицы, на картофельном поле только виргинильные и генеративные растения.
Ключевые слова:
щирица запрокинутая, онтогенез, онтогенетическая структура, динамика, однолетнее растение, инвазионный вид, онтогенетический спектр, онтогенетическое состояние, ценопопуляция, плотность
Abstract: The object of the study is green amaranth (Amaránthus retrofléxus L.) - a North American species, an invasive species for Russia, an annual weed. The aim of this work is to study the ontogeny and ontogenetic structure of the coenopopulations of the upturned squid. The research was carried out in 2014 in the vicinity of the village of Sizner, Mari-Turek region of the Republic of Mari El, in potato and beet fields. Within each field, 10-15 counting plots of 1 m2 in size were laid, where weekly observations of the development of beetle plants were carried out. To assess the density and ontogenetic structure of populations, we analyzed the R × C contingency tables using Fisher's exact test and the χ2 test (Glotov, 1984). The main conclusions of the conducted research include the following. For the first time, five ontogenetic states of the thrown-back beetle were identified (from seedlings to generative ones). The plant density of the squid in both fields does not differ and averages 6.9 ind./m2. The change in the ontogenetic structure of the coenopopulations of the upturned beetle in the studied fields occurs in different ways. The analysis showed that in the beet field, the plants undergo ontogenesis faster. In mid-July, on the beet field, all the ontogenetic states of the squid are presented, on the potato field, only virginal and generative plants.
Keywords: amaranth upturned, ontogenesis, ontogenetic structure, dynamics, annual plant, invasive species, ontogenetic spectrum, ontogenetic state, coenopopulation, density
Введение. Внедрение (инвазия) агрессивных чужеродных видов растений является одной из составных частей различных изменений биологического разнообразия и экологической значимости растительных сообществ [1, 2]. Выявление инвазионных видов является одной из актуальных задач изучения региональных флор [3].
Щирица (амарант) запрокинутая (Amaránthus retrofléxusL.) – североамериканский вид, широко расселившийся по всем континентам, является инвазионным видом, который включен в список видов растений «Черной книги флоры Средней России» [4]. Это распространённый, весьма агрессивный сорняк, одним из первых появляется на пустырях и заброшенных сельхозугодиях. Предпочитает гумусные, водопроницаемые, богатые питательными веществами, а прежде всего азотом, почвы [5]. Произрастает почти во всех районах России, в том числе обычен во всех областях Средней России.
Первое упоминание о щирице запрокинутой в Республике Марий Эл встречается в исследованиях Б. Василькова [6]. В настоящее время щирица запрокинутая встречается часто по всей территории республики, на полях, огородах, сорных местах близ жилья [7].
Цель данной работы – изучение онтогенеза и динамики онтогенетической структуры ценопопуляций щирицы запрокинутой.
Методика исследования. Исследования проводились в 2014 г. в окрестностях деревни Сизнерь Мари-Турекского района Республики Марий Эл на картофельном (ценопопуляция ЦП 1) и свекольном (ЦП 2) полях. Флористический состав сорных растений исследуемых полей сходный. На обоих полях были обнаружены щирица запрокинутая AmaranthusretroflexusL., вьюнок полевой Convolvulusarvensis L. и ромашка душистая Matricaria recutita L. На свекольном поле также встречалась лебеда лоснящаяся Atriplex sagittata Borkh.
В пределах каждого поля было заложено по 10-15 учетных площадок размером 1м2, на которых велись наблюдения за динамикой развития растений щирицы. Для этого еженедельно (2 июня, 9 июня, 16 июня, 23 июня, 30 июня, 7 июля, 14 июля) подсчитывали все растения щирицы на учетных площадках, определяли онтогенетическое состояние.
Для описания онтогенетических состояний щирицы использовалась классификация, разработанная Т. А. Работновым [8, 9] и дополненная А. А. Урановым [10, 11] и его учениками [12].
Для оценки плотности и онтогенетической структуры растений на учетных площадках в разные сроки наблюдения проводили анализ таблиц сопряженности R×C с помощью точного критерия Фишера [13] (в программе RCEXACT) и критерия χ2 [14].
Результаты исследования.
Онтогенез щирицы запрокинутой
Нами описан онтогенез щирицы (рис. 1). Описание проростков дано по литературным источникам.
Проростки (p) – ортотропные однопобеговые растения высотой до 2,5 см с двумя постепенно суженные в черешок семядолями. Появляется первый настоящий лист. Семядольные листья сверху серовато-зеленые, снизу фиолето-красные, длиной 0,7-0,9 см, шириной 0,1-0,3 см. Гипокотиль грязно-малиновый [15]. Главный корень имеет длину 1-3 см, боковых корней нет.
Ювенильные растения (j) – ортотропные однопобеговые растения высотой 2-4 см, имеют 2-4 хорошо развитых настоящих листа. Первая пара листьев округлой широко-яйцевидной или ромбическо-округлой формы с выемчатой верхушкой. Вторая пара листьев округло-ромбической или яйцевидной формы с суженным основанием и притупленной или слегка выемчатой верхушкой. Главная жилка заканчивается маленьким острием. Число боковых жилок на листовой пластинке 7-9. Длина листьев 1,0-1,5 см, шириной 0,6-0,9 см. Могут сохраняться семядольные листья. Эпикотиль короткий, мелковолосистый, 0,8-1,2 см длиной. Гипокотиль грязно-малиновый. Хорошо выражен главный корень длиной 1,5-4 см, боковые корни до 3 мм длиной. При повреждении главного корня 1-3 боковых корня I порядка достигают длины 1-2 см.
Имматурное растение (im) – ортотропное однопобеговое растение высотой 3-10 см с 4-9 хорошо развитыми настоящими листьями и 1-2 не развернувшимися листьями. Листья на длинных черешках, яйцевидные, яйцевидно-продолговатые или ромбические с клиновидным основанием. Длина листовой пластинки 1,7-4,3 см, шириной 1,4-2,8 см. Число боковых жилок 10-12. Хорошо выражены первые четыре междоузлия. Главный корень хорошо выражен, длиной до 7 см, имеются боковые корни I и II порядков ветвления длиной 1,5-2,0 см.
Виргинильное растение (v) – ортотропное однопобеговое растение, высотой 15-17 см. Имеется до 14 настоящих листьев с ромбической цельной пластинкой. Листья короткочерешковые, яйцевидно-продолговатые или ромбические с клиновидным основанием. Длина настоящего листа 0,7-5,3 см. Число боковых жилок 13. Основание листовой пластинки суженное. Лист сильно опушен с верхней и нижней стороны листа. Появляются боковые побеги, т.е. начинается ветвление. Главный корень имеет длину 7,5-10 см, формируются боковые корни I порядка.
Генеративное растение (g) – ортотропное однопобеговое или ветвистое растение высотой 20-30 см или более. Листья длиной 0,6-8,2 см, короткочерешковые, яйцевидно-продолговатые или ромбические с клиновидным основанием, основание листовой пластинки суженное, листовая пластинка опушена. Число боковых жилок 17. Формируется верхушечное соцветие зеленого цвета размером 6-10 см, имеются боковые соцветия. Главный корень имеет длину 11,5-15,0 см, формируются боковые корни I-IVпорядков.
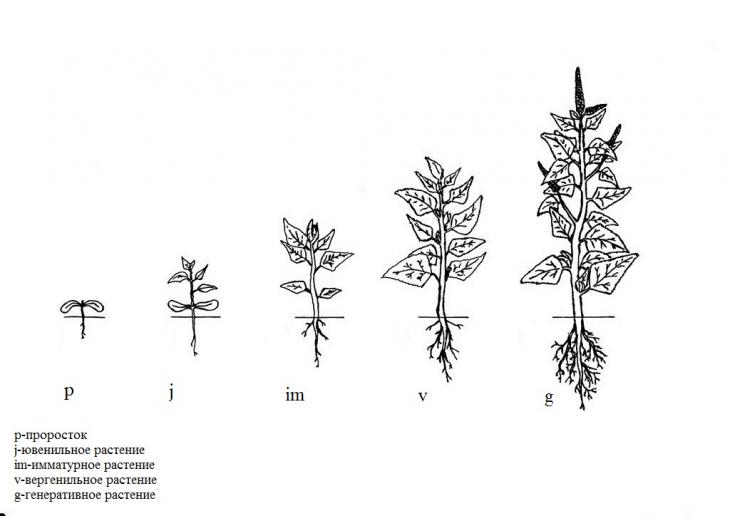
Рисунок 1 – Онтогенез щирицы запрокинутой
Динамика структуры ценопопуляций. Плотность растений щирицы на обоих полях не различается (Р>0,05) и составляет в среднем 6,9 экз./м2. Нами была изучена динамика онтогенетической структуры ценопопуляций щирицы на исследуемых полях. Анализ показал, что в пределах поля распределения по онтогенетическим состояниям между учетными площадками в первый (02.06.2014) и последний (14.07.2014) сроки наблюдения однородные (Р>0,13). В остальные сроки (с 09.06.2014 по 07.07.2014) распределения по онтогенетическим состояниям на разных учетных площадках в пределах поля различные (Р˂0,00001). Число растений, зарегистрированных на учетных площадках в разные сроки наблюдения, варьирует от 63 до 102 экземпляров на картофельном поле, от 43 до 103 экземпляров на свекольном поле.
В первый срок (02.06.2014) на обоих полях появляются только проростки. Через неделю (09.06.2014) часть растений переходит в ювенильное онтогенетическое состояние, а также появляются проростки щирицы. Во второй срок наблюдения распределения по онтогенетическим состояниям между полями однородное (P=0,04) (рис. 2).
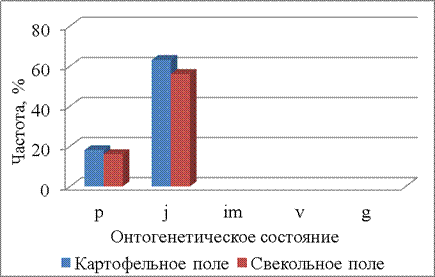
|
Рисунок 2 – Онтогенетические спектры щирицы запрокинутой в разных местообитаниях (09.06.14)
|
В третий срок наблюдения (16.06.2014), т.е. уже во второй декаде июня появляются растения имматурного онтогенетического состояния, а на свекольном поле часть растений переходит в виргинильное онтогенетическое состояние. Онтогенетические спектры (рис. 3) на двух полях в этот срок наблюдения различаются (P=0,000001). На свекольном поле в третий срок учета представлены все онтогенетические состояния от проростков до виргинильных, причем доля имматурных и виргинильных растений составляет более 40%. На картофельном поле наибольшая частота приходится на имматурные растения – около 60%.
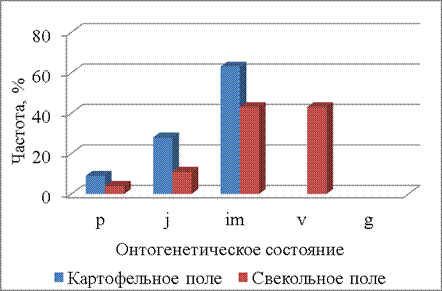
Рисунок 3 – Онтогенетические спектры щирицы запрокинутой в разных местообитаниях (16.06.2014)
В четвертый срок наблюдения (23.06.2014) на обоих полях представлены все онтогенетические состояния от проростков до виргинильных растений (рис. 4). Онтогенетические спектры различаются (Р=0,002). Частота имматурных и виргинильных растений более 30%. На обоих полях появляются проростки, но их частота на свекольном поле выше (более 10%), чем на картофельном (менее 3%).
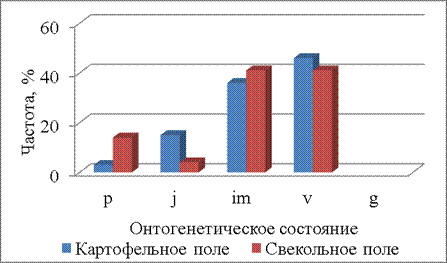
Рисунок 4 – Онтогенетические спектры щирицы запрокинутой в разных местообитаниях (23.06.2014)
К концу июня (30.06.2014) на обоих полях появляются генеративные растения (рис. 5). Однако на картофельном поле частота таких растений небольшая (менее 10%), а на свекольном поле составляет более 40%. Т.е. на свекольном поле растения щирицы быстрее проходят свой онтогенез, а на картофельном поле происходит задержка развития растений в прегенеративном периоде. На картофельном поле наибольшая частота растений приходится на виргинильное онтогенетическое состояние (более 60%), проростки отсутствуют, частота ювенильных и имматурных растений невелика – не более 15%. Онтогенетические спектры на картофельном и свекольном полях различны (Р=0,000001).
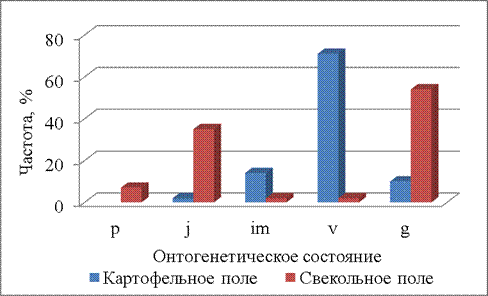
Рисунок 5 – Онтогенетические спектры щирицы запрокинутой в разных местообитаниях (30.06.2014)
В начале июля (07.07.2014) онтогенетическая структура на двух полях также различна (Р=0,000001). По сравнению с предыдущим сроком на свекольном поле увеличивается доля генеративных растений (до 70%), на картофельном поле она не изменяется (рис. 6).
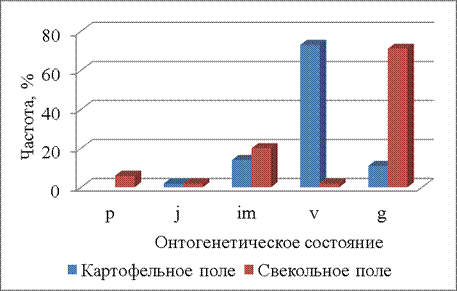
Рисунок 6 – Онтогенетические спектры щирицы запрокинутой в разных местообитаниях (07.07.2014)
В середине июля (14.07.2014) практически все растения переходят в генеративное онтогенетическое состояние (рис. 7). На свекольном поле в это время еще обнаружены проростки, ювенильные, имматурные и виргинильные растения, но их доля не превышает 5%. На картофельном поле из прегенеративного периода встречаются только виргинильные растения.
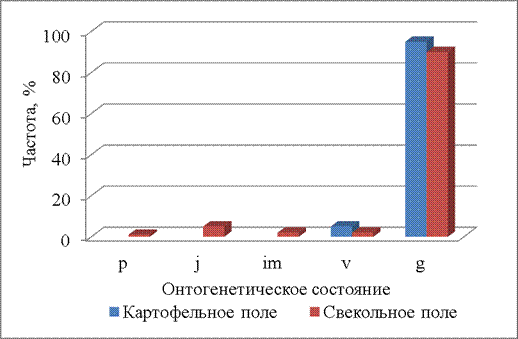
Рисунок 7 – Онтогенетические спектры щирицы запрокинутой в разных местообитаниях (14.07.2014)
Таким образом, изменение онтогенетической структуры ценопопуляций щирицы запрокинутой на исследуемых полях происходит по-разному. На свекольном поле растения проходят онтогенез быстрее, они быстрее переходят в генеративное состояние. Также на свекольном поле более растянуто по времени появление всходов – мы наблюдаем их еще в середине июля, когда большая часть растений достигли генеративного периода. В то же время на картофельном поле проростки наблюдаются только до 23 июня.
Однако нужно иметь в виду что у однолетних растений выявляется сезонная сменяемость онтогенетических спектров, когда в начале сезона в популяциях преобладают прегенеративные растения, в середине – генеративные, а к осени – генеративные и сенильные. Ценопопуляции однолетних растений иногда называют ложноинвазионными популяциями. Это связано с тем, что популяции однолетних сорных растений в агрофитоценоаах искусственно поддерживаются, когда после дождей или после уборки урожая массово появляются всходы этих сорняков, но их быстро уничтожают культивацией посева или гербицидами. В отдельные благоприятные для них годы происходят массовые вспышки численности всходов таких растений, но они быстро отмирают, не переходя во взрослое или тем более в генеративное состояние [16]. Поэтому в зависимости от погодных условий онтогенетическая структура ценопопуляций однолетних растений в разные годы может существенно варьировать.
Выводы. На основании вышеизложенного можно сделать следующие выводы.
1. Впервые для щирицы запрокинутой выделено и описано пять онтогенетических состояний: от проростков до генеративных растений.
2. Онтогенетические спектры на картофельном и свекольном полях в разные сроки наблюдения различаются. На свекольном поле онтогенез растений проходит быстрее, чем на картофельном. В середине июля на свекольном поле представлены все онтогенетические состояния щирицы, на картофельном поле только виргинильные и генеративные растения.
Библиография
1. Папченков, В.Г. Флористические находки в бассейне Средней Волги / В.Г. Папченков // Ботанический журнал. – 2007. – Т. 92, № 10. – С. 1580-1588.
2. Тремасова, Н.А. Инвазионные виды растений Ярославской области / Н.А. Тремасова, М.А. Борисова, Е.А. Борисова // Ярославский педагогический вестник. – 2012. – Т. 3, № 1. – С. 103-111.
3. Соколова, И.Г. Инвазивные виды Псковской области / И.Г. Соколова //Псковский регионологический журнал. – 2012. – № 14. – С. 97-102.
4. Виноградова, Ю.К. Черная книга флоры Средней России (Чужеродные виды растений в экосистемах Средней России) / Ю.К. Виноградова, С.Р. Майоров, Л.B. Хорун. – М.: ГЕОС, 2009. – 494 с.
5. Губанов, И. А. Иллюстрированный определитель растений Средней России / И.А. Губанов. – М., 2003. – 672 с.
6. Васильков, Б.П. К истории флоры Марийской области. (Из работ геоботанической экспедиции Марийского научно-исследовательского института) / Б.П. Васильков // М.А.О. (Марийская автономная область). – 1933. – № 11-12. – С. 71-87.
7. Абрамов, Н.В. Конспект флоры Республики Марий Эл / Н.В. Абрамов. – Йошкар-Ола: МарГУ, 1997. – 192 с.
8. Работнов, Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах / Т.А. Работнов // Труды БИН АН СССР. – Сер. 3. Геоботаника. – 1950. – Вып. 6. – С. 7-204.
9. Работнов, Т.А. Вопросы изучения состава популяций для целей фитоценологии /Т.А. Работнов // Проблемы ботаники. – Т. 1. – 1950. – С. 465-483.
10. Уранов, А.А. Онтогенез и возрастной состав популяций (вместо предисловия) / А.А. Уранов // Онтогенез и возрастной состав популяций цветковых растений. – М.: Наука, 1967. – С. 3-8.
11. Уранов, А.А. Возрастной спектр фитоценопопуляции как функция времени и энергетических волновых процессов / А.А. Уранов // Научные доклады высшей школы. Биол. науки. – 1975. – № 2. – С. 7-34.
12. Ценопопуляции растений (основные понятия и структура). – М.: Наука, 1976. – 277 с.
13. Хромов-Борисов, Н.Н. Биометрические задачи в популяционных исследованиях / Н.Н. Хромов-Борисов, Г.Б. Лаззаротто, Т.Б. Кист // Методы популяционной биологии. Матер. докл. VII Всерос. популяционного семинара. – Сыктывкар, 2004. – С. 62-86.
14. Глотов, Н.В. Биометрия / Н.В. Глотов, Л.А. Животовский, Н.В.Хованов, Н.Н. Хромов-Борисов. – Л.:ЛГУ, 1982. – 352 с.
15. Фисюнов, А.В. Определитель всходов сорных растений / А.В. Фисюнов. – Киев: Урожай, 1976. – 227 с.
16. Злобин, Ю. А.. Популяционная экология растений; современное состояние, точки роста; монография. – Сумы: Университетская книга, 2009. – 263 с.
References
1. Papchenkov, V.G. Floristicheskie nakhodki v basseine Srednei Volgi / V.G. Papchenkov // Botanicheskii zhurnal. – 2007. – T. 92, № 10. – S. 1580-1588.
2. Tremasova, N.A. Invazionnye vidy rastenii Yaroslavskoi oblasti / N.A. Tremasova, M.A. Borisova, E.A. Borisova // Yaroslavskii pedagogicheskii vestnik. – 2012. – T. 3, № 1. – S. 103-111.
3. Sokolova, I.G. Invazivnye vidy Pskovskoi oblasti / I.G. Sokolova //Pskovskii regionologicheskii zhurnal. – 2012. – № 14. – S. 97-102.
4. Vinogradova, Yu.K. Chernaya kniga flory Srednei Rossii (Chuzherodnye vidy rastenii v ekosistemakh Srednei Rossii) / Yu.K. Vinogradova, S.R. Maiorov, L.B. Khorun. – M.: GEOS, 2009. – 494 s.
5. Gubanov, I. A. Illyustrirovannyi opredelitel' rastenii Srednei Rossii / I.A. Gubanov. – M., 2003. – 672 s.
6. Vasil'kov, B.P. K istorii flory Mariiskoi oblasti. (Iz rabot geobotanicheskoi ekspeditsii Mariiskogo nauchno-issledovatel'skogo instituta) / B.P. Vasil'kov // M.A.O. (Mariiskaya avtonomnaya oblast'). – 1933. – № 11-12. – S. 71-87.
7. Abramov, N.V. Konspekt flory Respubliki Marii El / N.V. Abramov. – Ioshkar-Ola: MarGU, 1997. – 192 s.
8. Rabotnov, T.A. Zhiznennyi tsikl mnogoletnikh travyanistykh rastenii v lugovykh tsenozakh / T.A. Rabotnov // Trudy BIN AN SSSR. – Ser. 3. Geobotanika. – 1950. – Vyp. 6. – S. 7-204.
9. Rabotnov, T.A. Voprosy izucheniya sostava populyatsii dlya tselei fitotsenologii /T.A. Rabotnov // Problemy botaniki. – T. 1. – 1950. – S. 465-483.
10. Uranov, A.A. Ontogenez i vozrastnoi sostav populyatsii (vmesto predisloviya) / A.A. Uranov // Ontogenez i vozrastnoi sostav populyatsii tsvetkovykh rastenii. – M.: Nauka, 1967. – S. 3-8.
11. Uranov, A.A. Vozrastnoi spektr fitotsenopopulyatsii kak funktsiya vremeni i energeticheskikh volnovykh protsessov / A.A. Uranov // Nauchnye doklady vysshei shkoly. Biol. nauki. – 1975. – № 2. – S. 7-34.
12. Tsenopopulyatsii rastenii (osnovnye ponyatiya i struktura). – M.: Nauka, 1976. – 277 s.
13. Khromov-Borisov, N.N. Biometricheskie zadachi v populyatsionnykh issledovaniyakh / N.N. Khromov-Borisov, G.B. Lazzarotto, T.B. Kist // Metody populyatsionnoi biologii. Mater. dokl. VII Vseros. populyatsionnogo seminara. – Syktyvkar, 2004. – S. 62-86.
14. Glotov, N.V. Biometriya / N.V. Glotov, L.A. Zhivotovskii, N.V.Khovanov, N.N. Khromov-Borisov. – L.:LGU, 1982. – 352 s.
15. Fisyunov, A.V. Opredelitel' vskhodov sornykh rastenii / A.V. Fisyunov. – Kiev: Urozhai, 1976. – 227 s.
16. Zlobin, Yu. A.. Populyatsionnaya ekologiya rastenii; sovremennoe sostoyanie, tochki rosta; monografiya. – Sumy: Universitetskaya kniga, 2009. – 263 s.
Результаты процедуры рецензирования статьи
В связи с политикой двойного слепого рецензирования личность рецензента не раскрывается.
Со списком рецензентов издательства можно ознакомиться здесь.
Предмет исследования: является, по мнению автора статьи изучение особенности онтогенеза и динамики онтогенетической структуры ценопопуляций щирицы запрокинутой. Для успешного применения мероприятий по борьбе с сорняком необходимо знать особенности семенного размножения в условиях интродукции. Однако давая характеристику щирица как злостному сорняку, автор статьи упускает из виду том, что это очень прогрессивный с точки зрения селекцией вид растения, нашедший в последнее время широкое распространение как поставщик высококачественного белка и масла. Используя название щирица автору статьи следовало всё-таки заменить его на амарант.
Методология исследования: Авторы позиционируют как измерение встречаемости в флористическом составе сорных растений различных онтогенетических стадий. Однако, на наш взгляд, было выявить типа ценотические отношения: конкуренции или симбиоза, а возможно и квартирарство.
Актуальность проблемы состоит в выявлении в том, что сорные растения являются исходной базой для селекционной работы, декоративными и лекарственными растениями и для их успешного семенного размножения необходимо знать показатели онтогенеза в условиях интродукции к изменениям климата с позиции повышения эффективности природопользования.
Научная новизну автор статьи не позиционирует, ими проведен анализ литературных данных по характеристики и оценки в условиях калины для их сроков онтогенеза и встречаемости в условиях разного флористического состава, однако они забывают об особенностях агротехники и полива.
Стиль, структура, содержание стиль изложения результатов научный, структура построения и изложения материала выстроена достаточно логично. Однако есть ряд вопросов, в частности: автору статьи следовало бы сделать вступление, введение сильно перегружено справочным материалом, однако не полон в связи с оценкой рода этого растения и перспектив его использования. На наш взгляд является излишним перечисление особенностей объектов исследования с точки зрения достаточно пространной цитаты. , Авторами на основе данных оставлены таблицы, которые носят характер визуализации текстовой информации в графической форме, при этом отсутствует анализ разнородности состава по мере прохождения стадий цикла развития.
Статья изобилует ссылками и цитатами, что придает ей характер реферативного литературного обзора.
Библиография обширна, содержит в основном классические источники.
Апелляция к оппонентам в выявлении проблемы на уровне имеющейся информации, полученной автором в результате анализа ограниченного ряда литературных источников.
Выводы, интерес читательской аудитории Приводимый автором вывод резюмирует содержание статьи, выводы статьи является констатирующими, отсутствует объяснения причин почему происходят разные сроки прохождение стадии развития на картофельном и свекольном полях. Среди возможных причин таких как конкуренция питательные вещества, или особенностей внесение удобрений, или особенности увлажнения авторами не рассмотрена не одна из причин. а потребитель представленной информации в статье авторами не определён.
|








