|
DOI: 10.7256/2453-8922.2017.2.22777
Дата направления статьи в редакцию:
23-04-2017
Дата публикации:
24-06-2017
Аннотация:
Объектом исследования данной статьи являются полигольнально-жильные структуры, сформировавшиеся в течение голоцена в пределах зоны современных арктических тундр на территории острова Белый и в долине реки Тамбей на северо-востоке полуострова Ямал. Исследованы палиноспектры повторно-жильных льдов и вмещающих их отложений, а также их химический состав. Для обоснования палеогеографических реконструкций проанализированы как имеющиеся у автора радиоуглеродные датировки, так и полученные другими исследователями из голоценовых отложений на севере Полуострова Ямал и близлежащих территоорий Основной метод исследования - палинологический. Обработка образцов из вмещающих отложений проводилась по стандартной методике, но без использования жидкости Эрдтмана, из повторно-жильных льдов концентрат пыльцы и спор извлекался по упрощенной методике из-за низкой концентрации пыльцы и спор . Основным результатом проведенного исследования является систематизация данных палинологического анализа голоценовых повторно-жильных льдов, находящихся в зоне воздействия изменений уровня моря. Показано, что повторно-жильные льды в долине реки Тамбей накапливались при участии не только атмосферных осадков, но и в режиме подтопления морскими водами. Повторно-жильные льды в толще первой морской террасы на острове Белый характеризуются аномально высоким содержанием для повторно-жильных льдов морских солей, однако, это засоление не связано с изменениями уровня моря, а появилось вследствие накопления повторно-жильных льдов на берегах небольших мелководных остаточных озер, с высокой соленостью.
Ключевые слова:
спорово-пыльцевые диаграммы, локальные палинозоны, полуостров Ямал, радиоуглеродный возраст, ледяные жилы, палиноспектры, голоцен, торфяник, климатичнские колебания, Север Западной Сибири
УДК: 551.89
Работа частично финансируется грантом РФФИ 17-05-00794
Abstract: Holocene ice wedge structures in the arctic tundra in the Tambey river valley and in the Belyi Island are the subject of the study. The author studies the pollen spectra in the ice wedges and surrounding sediments and their chemical composition. To substantiate paleograhic reconstructions, the author analyzes radiocarbon datings of the Holocene sediments from the studied areas and the surrounding territories. Pollen analysis is the main research method. Adjacent sediment samples have been studied according to the standard procedure, but without acetolysis mixture. Pollen has been extracted using the simplified method due to low pollen concentration. The main result of the study is the systemization of the data of palynological analysis of Holocene ice wedges located in the zone of sea-level changes' imact. The author shows that ice wedges in the Tambey River valley had been accumulated from precipitations and because of the inundation by marine water. Ice wedges in the first marine terrace of the Belyi Island are characterized by the abnormally high concentration of marine salt, but the salinization is caused by ice wedge formation on the sides of remnant shallow saline lakes, not by sea-level changes.
Keywords: sporo-pollen charts, local pollen zone, Yamal Peninsula, radiocarbon age, ice wedge, pollen spectra, Holocene, peat, climatic fluctuations, North-West Siberia
ВВЕДЕНИЕ
Ландшафты Арктики при всей своей простоте обладают своеобразной внутренней структурой. Живые организмы находятся в экстремальных условиях обитания, температурный фактор- основное экологическое ограничение, поэтому влияние температурных изменений на связи между отдельными группами живых организмов здесь гораздо выше. Даже незначительные изменения условий могут приводить к заметным изменениям растительного покрова, почв и других компонентов ландшафта.
Объектом исследования данной статьи является история развития растителдьного покрова полигонально-жильных структур, сформировавшихся в течение голоцена в пределах подзоны современных арктических тундр на территории острова Белый 73°14'39.1"с.ш., 70°13'16.4" в.д. и в долине реки Тамбей (71°41'54.63" с.ш., 71°24'29.95" в.д.) на северо-востоке полуострова Ямал. Эти точки расположены на западе и востоке подзоны арктических тундр. К подзоне арктических тундр относится северная оконечность полуострова, примерно до 71°20' с.ш. [1]. Если долина реки Тамбей, как и все восточное побережье полуострова находится под воздействием Обской губы, то на о. Белый, особенно его на западное побережье воздействует Карское море (рис. 1).
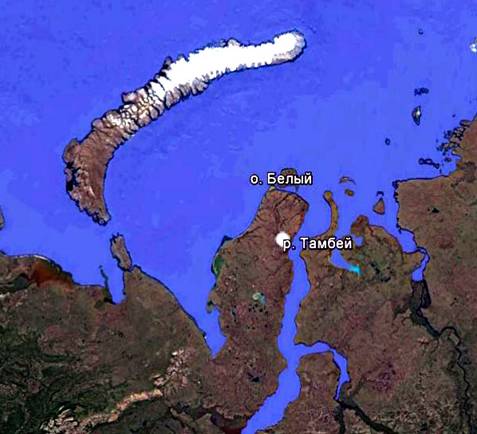
Рис. 1. Местоположение палинологически изученных голоценовых полигонально-жильных структур на острове Белый и на пойме р.Тамбей.
Голоценовые повторно-жильные льды широко распространены и на острове Белый и на низких элементах рельефа - на поймах рек и озер близ пос. Тамбей на севере Ямала. В пределах этих территорий преобладает аккумулятивный тип рельефа, который представлен различными по возрасту и генезису геоморфологическими уровнями. В районе долины р.Тамбей выделяются среднеплейстоценовая и позднеплейстоценовая морские равнины, комплексы позднеплейстоценовых морских, лагунно-морских, озерных и аллювиальных террас, голоценовая пойма и лайда, на о.Белом выделяется комплекс морских террас и лайда [2]. Всюду развиты песчаные раздувы.
Средняя годовая температура воздуха на этой территории -10…-11 °С. Средние январские изотермы имеют меридиональное, изменяясь от -22 °С на западе до -28 °С на востоке зоны. Средняя июльская температура воздуха в арктической тундре возрастает с северо-запада (4 °С) на юго-восток (10 °С). Условия для зимовки растений суровые — средний из абсолютных минимумов температуры воздуха за зиму (год) составляет -48…-43 °. Продолжительность безморозного периода в арктической тундре колеблется в пределах 49-64 дней, сумма положительных температур - менее 707 °С (а выше 5 °С - менее 614 °С), что является существенным ограничением прироста биомассы в теплый период года (http://www.activestudy.info/klimat-arkticheskoj-tundry). Режим увлажнения в тундре - избыточный. Количество осадков составляет 250-310 мм за год и возрастает с севера на юг. Установление снежного покрова отмечается в начале октября. Продолжительная и холодная зима благоприятствует накоплению снега, но сильные зимние ветры сдувают его. Высота снежного покрова составляет всего 28-43 см. Весной разрушение устойчивого снежного покрова заканчивается в арктической тундре к концу июня.
Ветровой режим в приземном слое воздуха характеризуется высокой повторяемостью сильных ветров. Средние за год скорости ветра составляют 6-9 м/с и наибольшие на побережье. Здесь отмечается максимум повторяемостей сильных ветров (15 м/с и более) - более 50 дней. Зимой и в переходные сезоны преобладают юго-западные ветры (40- 65 % случаев), а летом — северные. (http://www.activestudy.info/klimat-arkticheskoj-tundry). Ветровой режим играет важную роль для формирования палиноспектров в пределах таких открытых ландшафтов как тундра. Из-за преобладания юго-западных ветров весной, на поверхность выпадает пыльца преимущественно древесных пород, произрастающих за сотни километров к юго-западу. Преобладание северных ветров в летний сезон приводит к тому что в летних палиноспектрах представлены в основном тундровые виды растений.
В подзоне арктических тундр на водоразделах в основном встречаются следующие полидоминантные фитоценозы: лишайниково-моховые, мохово-лишайниковые, кустарничково-моховые и кустарничково-лишайниковые. Участие травянистых растений злаков, осок и разнотравья варьирует от спорадического до значительного. Весьма существенную роль в структуре растительного покрова играют травяно-моховые тундры и травяно-моховые болота. В речных долинах и на месте спущенных озер формируются лугоподобные сообщества [3].
В низовьях реки Тамбей растительность представлена различными видами лишайников, главным образом кладонией [4], мхов, тундровых трав в основном пушицы узколистной и Шейхцера, разных видов осок, местами - стелющейся карликовой ивой. Распространены осоково-гипновые полигональные болота на торфяно-глеевых почвах.
На о. Белом распространены приморские заливные луга и мелкокочковатые моховые тундры (рис. 2) с гипновыми мхами, среди которых присутствуют лишайники (кладония, пельтигера). Травяной покров состоит главным образом из осок пушиц узколистной и Шейхцера и злаков: дюпонтии Фишера, зубровки редкоцветной. Заросли арктофилы рыжеватой окаймляют понижения, приуроченные обычно к берегам термокарстовых озёр [5].
Рис. 2. Растительность острова Белый: а - в -терраса, г - небольшое озеро, д-е - приморские заливные галофитные луга (тампы). Фото с сайтов http://two-hitchhikers.ru/priroda-arkticheskogo-ostrova-belyiy/ и http://severok1979.livejournal.com/95859.html
В последние годы исследователями отмечается тенденция к «позеленению» Арктики [6], хотя на территории арктических тундр эта тенденция проявляется не так заметно, как, например, в северной лесотундре. Тем не менее появление ольховника отмечается в растительных сообществах на плакорах, там, где раньше его не было, увеличивается также роль травянистых сообществ. По сути в настоящий момент мы наблюдаем смену одного стабильного состояния на другое. Подобные изменения происходили и ранее, о чем свидетельствуют палинологические данные. Изучение повторно-жильных льдов позволило зафиксировать колебания в составе пыльцевого дождя в голоцене.
МЕТОДЫ
Основной метод исследования – палинологический, дополнительно использованы результаты химического и анализа повторно-жильных льдов и данные по 14С датированию голоценовых многолетнемерзлых отложений в изученных районах. Для решения поставленной задачи- реконструкции истории развития полигольнально-жильных структур, были палинологически изучены как повторно-жильные льды, так и вмещающие их отложения. В качестве основы для интерпретации использованы результаты изучения поверхностных проб и разных растительных подзон Ямала и разных фациальных обстановок [7]. В процессе палинологического анализа использовались эталоны пыльцы современной растительности Ямала. Особое внимание уделено морфологии пыльцевых зерен карликовой березки.
РЕЗУЛЬТАТЫ
Палинологическая характеристика субфоссильных спектров
При интерпретации палинологических данных в Арктике и Субарктике криогенный фактор практически не учитывается, в то время как многие особенности палиноспектров обусловлены формированием в холодном климате в зоне многолетнемерзлых пород. Решение проблемы интерпретации результатов палинологического анализа в зоне развития многолетнемерзлых пород складывается из решения частных вопросов. Оценка концентрации пыльцы и спор на территории тундровой зоны и исследование палиноспектров из повторно-жильных льдов - это обоснование дополнительных критериев оценки и интерпретации палиноспектров.
Для палиноспектров из отложений в пределах подзоны арктических тундр наиболее низкая концентрация характерна для поверхностных отложений пляжа и речных осушек, а наиболее высокая – для поверхности высокой поймы или лайды (табл. 1).
Таблица 1. Средние значения концентрации пыльцы экз./см2 в поверхностных пробах в Российской Арктике и Субарктике. По данным А.Васильчук [7]
|
Расти-
тельная подзона
|
Точки отбора
|
Водо-
разде-
лы
|
Низ-
кая
пой-
ма
|
Реч-
ные
осуш-
ки
|
Лай-
да
|
Пляж
|
|
Арктические тундры
|
Матюй-сале
|
7
|
18
|
4
|
20
|
8
|
|
О.Белый
|
5
|
14
|
3
|
22
|
16
|
|
р.Харасавэй
|
66
|
5
|
16
|
46
|
28
|
|
Гыда
|
54
|
81
|
7
|
63
|
39
|
|
Хатанга
|
32
|
–
|
–
|
–
|
–
|
Исследование субфоссильных палиноспектров отложений пляжа Карского моря и Обской губы показало, что современные палиноспектры в отложениях разного гранулометрического состава могут резко различаться, особенно по составу дальнезаносных компонентов. Это, на наш взгляд, связано с вариациями концентрации пыльцы и спор в отложениях пляжа, где пыльца и споры находятся под постоянным волновым воздействием (табл. 2). Содержание пыльцы и спор (%) в отложениях пляжа Карского моря и Обской губы показывает, что гидродинамические процессы и режим осадконакопления в первую очередь определяют особенности палиноспектров пляжевых фаций, а информация о составе окружающей растительности содержится или в неявном виде или вообще искажена. Между гранулометрическим составом пляжевых осадков и степенью отражения растительного покрова в палиноспектрах, как правило, прослеживается отчетливая зависимость – в грубозернистых разностях привнос древних и дальних палиноморф значительно выше, иногда они даже могут преобладать над современными пыльцой и спорами.
Проведенные исследования субфоссильных палиноспектров на о.Белом показали, что в их составе доминирует пыльца злаков и осок, которая в сумме составляет более 60%, отражая преобладание растений этих семейств в растительном покрове. Пыльца остальных встречающихся в растительном покрове семейств также отмечается, но, как правило, единично, часто не соответствуя их роли в фитоценозах. Для субфоссильных спектров характерно сравнительно высокое содержание заносной пыльцы сосны обыкновенной (до 10%) и сибирского кедра (до 13%), содержание пыльцы ели в большинстве изученных образцов составило около 2%. Очевидно, это фоновые региональные значения. Роль пыльцы кустарников карликовой березки и ольховника несущественна (1–4%), пыльца ивы встречена в единственной поверхностной пробе (непосредственно рядом с растением), пыльца полыни встречается единично.
В субфоссильных палиноспектрах в долине р.Тамбей доминирует пыльца осок, которая составляет около 5,2-16,8%, а также злаков (3,5-12%), что примерно соответствует участию этих семейств в растительном покрове. Пыльца иных растений, встречающихся в растительном покрове, отмечается, как правило, единично, часто не соответствуя их роли в растительных сообществах. Для субфоссильных спектров характерно сравнительно высокое содержание заносной пыльцы сосны обыкновенной (до 8-9%) и сибирского кедра (до 10-12%). Содержание пыльцы ели в большинстве изученных образцов составило менее 2%, карликовой березы - до 18%, ольховника (дюшекии) не более 5-8% [8]. Сравнивая состав поверхностных проб отметим, что характеристика субфоссильных палиноспектров очень близка. Вариации содержания пыльцы древесных пород, кустарников, а также основных групп травянистых растений злаков, осок, верескоцветных находятся в одном диапазоне, что очевидно соответствует подзоне арктических тундр. Различие отмечается для пыльцы кустарников, содержание пыльцы карликовой березки и ольховника в долине р.Тамбей несколько выше, что связано с болле южным расположением долины р.Тамбей.
Таблица 2. Содержание пыльцы и спор (%) в отложениях пляжа Карского моря и Обской губы
|
Точка отбора
|
о.Белый
|
факт.Матюй-сале
|
а/п МыКаменный
|
пос.Новый Порт
|
|
Зона
|
Арктическая тундра
|
Кустарничковая
тундра
|
Южные кустарниковые тундры
|
|
Литология
|
Мелкие пески и супеси
|
Крупные и средние пески
|
Мелкие пески и супеси
|
Крупные и средние пески
|
Мелкие пески и супеси
|
Крупные и средние пески
|
Мелкие пески и супеси
|
Крупные и средние пески
|
|
Пыльца деревьев
|
15.0
|
22.5
|
3.7
|
52.0
|
15.9
|
40.1
|
41.8
|
76.6
|
|
Пыльца кустарников
|
4.0
|
7.8
|
18.1
|
3.5
|
43.3
|
6.3
|
19.3
|
2.4
|
|
Пыльца трав и кустарничков
|
58.0
|
40.0
|
45.7
|
19.5
|
38.0
|
15.4
|
12.3
|
10.1
|
|
Споры
|
23.0
|
30.0
|
32.5
|
31.0
|
18.6
|
39.2
|
26.6
|
10.9
|
|
Larix
|
–
|
–
|
–
|
1.5
|
–
|
–
|
–
|
–
|
|
Pinus sibirica
|
13.9
|
9.2
|
2.9
|
28.0
|
4.0
|
12.4
|
1.5
|
14.7
|
|
Pinus sylvestris
|
–
|
10.0
|
0.8
|
15.5
|
5.3
|
10.2
|
0.6
|
23.0
|
|
Picea
|
–
|
2.5
|
–
|
2.0
|
2.7
|
4.3
|
1.2
|
16.6
|
|
Betula
|
1.1
|
0.8
|
–
|
4.5
|
4.0
|
9.6
|
30.8
|
13.1
|
|
Alnus
|
–
|
–
|
–
|
0.5
|
–
|
–
|
7.7
|
5.0
|
|
Betula sect. Nanae
|
1.0
|
4.2
|
3.7
|
3.0
|
14.1
|
3.9
|
15.4
|
2.4
|
|
Alnaster
|
–
|
1.7
|
8.0
|
0.5
|
10.6
|
2.4
|
1.2
|
0.6
|
|
Salix
|
3.0
|
1.7
|
6.4
|
–
|
18.6
|
0.1
|
2.7
|
–
|
|
Poaceae
|
10.8
|
25.8
|
6.4
|
0.5
|
15.0
|
4.2
|
4.5
|
1.1
|
|
Cyperaceae
|
10.1
|
5.0
|
23.8
|
4.5
|
1.8
|
4.2
|
1.5
|
3.2
|
|
Eryophorum
|
18.7
|
–
|
7.9
|
3.6
|
9.7
|
2.1
|
–
|
–
|
|
Ericaceae
|
11.3
|
1.7
|
6.7
|
4.5
|
2.7
|
0.7
|
0.3
|
1.1
|
|
Artemisia
|
–
|
3.3
|
–
|
1.0
|
5.7
|
–
|
0.3
|
0.3
|
|
Varia
|
1.0
|
4.1
|
2.4
|
0.5
|
2.9
|
4.2
|
1.8
|
2.3
|
|
Bryales
|
9.2
|
25.0
|
23.8
|
10.5
|
14.1
|
33.4
|
9.7
|
0.3
|
|
Sphagnum sp.
|
10.7
|
0.8
|
6.4
|
7.5
|
2.2
|
2.8
|
5.0
|
3.5
|
|
Equisetum
|
–
|
0.8
|
2.1
|
1.0
|
–
|
3.0
|
0.4
|
1.4
|
|
Polypodiaceae
|
–
|
–
|
–
|
3.0
|
1.8
|
–
|
5.9
|
0.6
|
|
Lycopodium sp
|
3.1
|
–
|
–
|
1.5
|
0.4
|
–
|
1.8
|
1.1
|
|
Hupezia selago
|
–
|
–
|
–
|
7.0
|
–
|
–
|
–
|
–
|
|
Экз/г
|
540
|
870
|
210
|
695
|
4822
|
416
|
982
|
602
|
|
Переотложенные дочетвертичные
|
2.7
|
29.2
|
37.0
|
30.5
|
23.2
|
26.1
|
28.2
|
18.1
|
| |
|
|
|
|
|
|
|
|
|

а
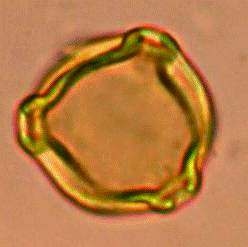
б
 . .
в
Рис. 3. Пыльцевые зерна Betula nana ssp. exilis. со слабо выраженными порами (а) и хорошо выраженными порами (б); варьирует также толщина экзины и размер пыльцевых зерен (в). Фото Л.А.Савельевой и Е.Рашке [9]
Из субфоссильных палиноспектров разных подзон тундр Западной Сибири проведены замеры диаметра пыльцевых зерен для 200 экземпляров пыльцевых зерен Betula nana L. (рис. 3). Размер пыльцевых зерен в поверхностных пробах варьировал существенно в пределах 15–33 μ. При этом для арктических тундр в точках в долине р.Тамбей и на о. Белом получено распределение. при котором практически все измеренные пыльцевые зерна попали в интервал 18–25 μ. Это означает, что для арктических тундр на территории полуострова Ямал пыльца карликовой березки является и региональным и дальнезаносным элементом. Очевидно, это связано. в основном. с повторяемостью ветров и их скоростью в открытых ландшафтах тундр. Концентрация пыльцы карликовой березки в поверхностных пробах арктических тундр составляет менее 10 экз./см2.

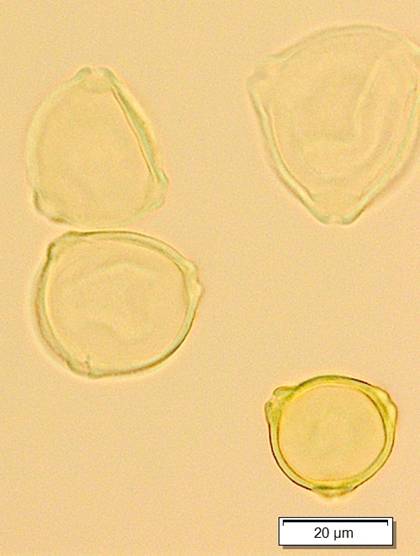
а

б
Рис. 4. Пыльцевые зерна Duschekia fruticosa (Alnaster fruticosus. Alnus viridis ssp. fruticosa) встречаются в основном со слабо выраженными порами. и практически незаметными арками (а). редко отмечаются более выраженные поры и арки (б) размеры пыльцевых зерен и толщина экзины варьируют. Фото Л.А.Савельевой и Е.Рашке [9]
Пыльца ольховника (дюшекии) важный компонент палиноспектров (рис. 4), поскольку в связи с повышением температур летнего сезона. которое отмечается в последние годы зафиксировано продвижение северной границы ареала этого растения, а также изменение состава травянистой растительности. увеличение разнообразия фитоценозов [6].
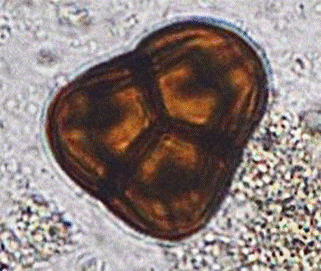
а

б
Рис. 5. Пыльцевые зерна Ledum palustre (а) и Vaccinium vitis-ideae (б). Фото Л.А.Савельевой и Е.А.Рашке [9]
Концентрация пыльцы верескоцветных (рис. 5) существенно меняется в разных тундровых экотопах и подзонах. У большинства видов верескоцветных максимальная продуктивность отмечается у границы тундры и лесотундры. Иногда концентрация пыльцы этих растений достигает значений 200-300 экз./см2. составляя в среднем 25-35 экз./см2. В поверхностных пробах на участках в долине р.Тамбей и на о.Белом концентрация составляет соответственно 2-4 экз./см2 и 1-2 экз./см2 соответственно.
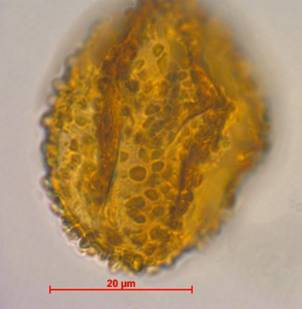
Рис. 6. Пыльцевое зерно Rubus Chamaemorus. Фото Л.А.Савельевой и Е.А.Рашке [9]
Пыльца Rubus Chamaemorus (морошки) в арктических тундрах (рис. 6) встречается постоянно. но в очень низкой концентрации 1-2 экз./см2. в фитоценозов арктических тундр пыльцевая продуктивность морошки очень низкая. Это компонент палиноспектров типичных (гипоарктических) тундр. появление пыльцы морошки в палиноспектре может свидетельствовать о развитии тундровых фитоценозов. оптимальная пыльцевая продуктивность этого растения отмечается в широком диапазоне от южных гипоарктических тундр до лесотундры.
Стратиграфия и криолитологическая характеристика разрезов
На о. Белом повторно-жильные структуры изучены в пределах плоской поверхности первой террасы. Поверхность характеризуется с выпуклыми полигонами размером от 5х5 до 7х10 м. В береговом обнажении рядом с метеостанцией вскрыта система двух сложно перекрещивающихся сингенетических повторно-жильных льдов. Вскрытая высота жил около 3 м (рис. 7). Ширина более узкой жилы в верхней части 0.46 м. ниже – до 0.65 м. ширина более широкой жилы – более 1 м. Лёд жилы прозрачный голубовато-серый. чётко вертикальнополосчатый. В верхней части насчитывается 34 элементарных жилки. а на глубине 1 м – 40 элементарных жилок. По обеим сторонам жилы выделяется “кайма” чистого стекловатого льда. шириной 3-4 см. На глубине 1 м справа от “каймы” ледогрунт представлен субвертикальными элементарными жилками. шириной 0.4-0.5 см. разделёнными вертикальными слоями песка. такой же ширины. В основном теле узкой жилы также встречены два ксенолита песка шириной до 1.5 см. а также небольшая прерывистая по вертикали жилка. сложенная опесчаненным торфом. шириной до 0.3-0.5 см. На глубине 1.5 м от этой ледяной жилы вправо отходит тонкая (шириной 4-5 см) ледяная жила. состоящая из 3 элементарных жилок. достигающая длины 0.9 м. В обнажении хорошо вскрыт контакт описанной выше узкой жилы и более широкой жилы. В теле широкой жилы этот контакт выделяется в виде шва, вдоль которого проходит полоса матового льда. шириной 2 см. основная часть разреза представлена песком мелким серым, с поверхности отмечены тонкие слои торфа и супеси.

б

Рис. 7. Обнажение отложений первой морской террасы острова Белый с сингенетическими повторно-жильными льдами. Фото Ю. Васильчука (а) и Л. Бойченко (б)
Из прослоя торфа в обнажении первой террасы из подошвы торфяника с глубины 2.5 м В.М.Леоновым получена радиоуглеродная датировка 8500 ± 120 лет (ЛУ-1139). Зарождение сильноминерализованных жил в отложениях первой морской террасы о. Белый, таким образом, датировано началом периода голоценового оптимума. Высокая минерализация льда жил (до 1240 мг/л) и преобладание в ионном составе хлора и натрия свидетельствует об участии морских вод в формировании жилы (рис. 8).
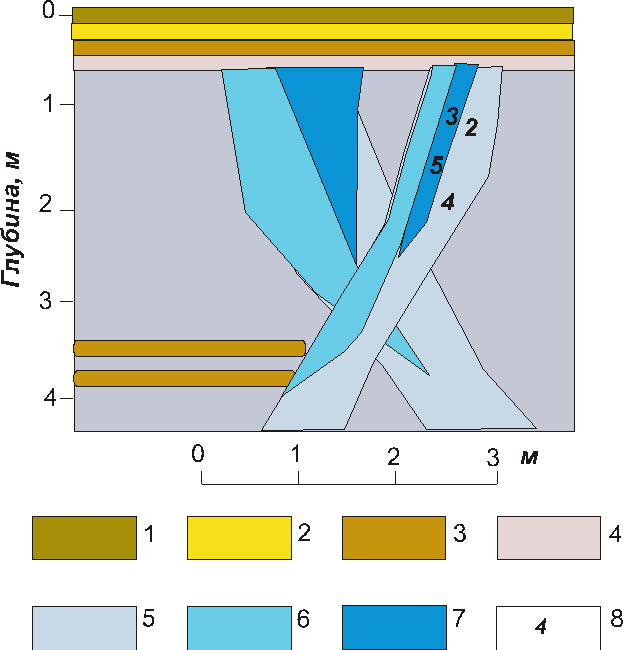
Рис. 8. Отбор образцов из повторно-жильных льдов в толще первой морской террасы на о. Белом: 1 – торф талый; 2 – песок мелкий серо-желтый талый; 3 – торф мерзлый 4 – песок мелкий серый мерзлый; 5 – точки отбора проб на палинологический и химический анализы; 6-8 – минерализация льда - мг/л: 6 – 400-500; 7 – 500-800; 8 – более 800
В долине р.Тамбей повторно-жильные структуры изучены в пределах высокой поймы. В 2 км севернее оз. Харато, были пробурены несколько скважин по створу от центра полигона до канавки и далее ряд зондировочных скважин по жиле для выявления ее боковых контактов и конфигурации ледяного клина (рис. 9). На поверхности поймы развит полигонально-валиковый рельеф с размерами полигонов от 10 х 10 м до 30 х 40 м. Разрез верхней части толщ поймы представлен песком русловой фации, залегающим в интервале 3.25-9.0 м от поверхности, перекрытым слоистой пачкой песков фации приречной поймы (мощностью 0,3 м); выше залегают пески лагунно-морской фации (мощностью 1,6 м), перекрытые супесью пойменной фации (0,15 м); разрез венчает песок аллювиальный (0,1 м) и торф (0,1-0,5 м). Песок, залегающий в основании разреза, светло-серого цвета, косослоистый, среднезернистый, с включениями мелких частичек угля и аллохтонного торфа. Криогенная текстура песка массивная, объемная льдистость 10-20‰. Очевидно, песок откладывался в начальную стадию аккумуляции аллювия. Приустьевое положение участка приводило к осолонению вод реки, что повлекло к отложению сравнительно сильно минерализованных русловых осадков. В обнажении поймы прослежена система ледяных жил (рис. 10). Их количество достигает 5-6 на каждые 85-100 м. Отмечены широкие и узкие ледяные жилы [10]. Строение узкой жилы двучленное. В правой половине жилы лед прозрачный. с обилием органического материала. в левой – молочно-белый. с отчетливо выраженной вертикальной слоистостью за счет элементарных жилок. Прозрачный лед характеризуется повышенным содержанием водорастворимых солей (до 0.013‰) гидрокарбонатно-хлоридного состава. с высоким. максимальным для всего комплекса. содержанием магния. Это позволяет сделать вывод о преимущественно субаквальном режиме формирования правой части жилы. а структурно-текстурные особенности льда – о значительном его метаморфизме. очевидно. обусловленном длительностью существования этой части льда. Молочно-белый лед в левой половине отличается меньший содержанием водно-растворимых солей (не более 0.008‰). что. вероятнее всего указывает на преимущественно субаэральный режим формирования льда.
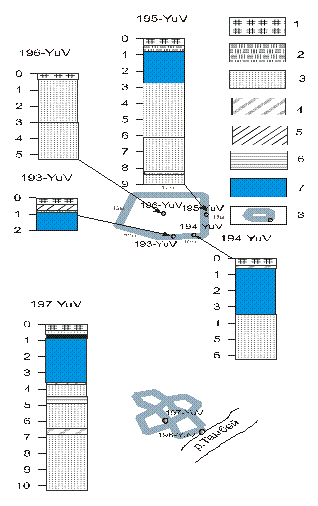
Рис. 9. Расположение и характеристика разрезов скважин (точки 193, 194,195,196,197-YuV), на высокой пойме реки Тамбей: 1 – торф талый; 2 – торф мерзлый льдистый; 3 – песок мелкий, мерзлый; 4 – супесь средняя мерзлая; 5 – суглинок тяжелый мерзлый; 6 – глина; 7 – повторно-жильный лед; 8 – расположение скважин в пределах полигонов по [10]

Рис. 10. Широкие (а) и узкие (б) голоценовые повторно-жильные льды с веерообразными изоклинальными складками на контактах, образованные маркирующим горизонтом слоистых оторфованных песков на участке поймы на левобережье в устье р.Тамбей, расположенном в 15 км северо-восточнее пос. Тамбей. Фото Ю.К. Васильчука
Палинологическая характеристика
Спорово-пыльцевые спектры песков в основании разреза высокой поймы ниже отметки 2,25 м (см. рис. 9) содержат большое количество пыльцы древесных пород с преобладанием пыльцы кедра (до 30%) и ели (до 25–28%). Довольно заметно о содержание пыльцы карликовой березы – до 30%. Такие палиноспектры нетипичны для отложений высокой поймы рек Восточного Ямала. Это позволяет предположить существование в период накопления частых нагонных явлений или подтопления устьевой части реки (исходя из повышенного содержания солей. преобладания ионов хлора и натрия и преобладания пыльцы сосны и ели которая переносится и по воздуху. и по воде). Высокое содержание пыльцы древесных пород позволяет отнести формирование отложений нижней части разреза высокой поймы к оптимуму голоцена. Слоистая пачка песка, залегающая в интервале 2,25–1,95 м характеризуется высокой объемной льдистостью – до 60‰ в верхней части горизонта. Наличие в слоистых песках прослоев торфа и относительно небольшое содержание водорастворимых солей (менее 0.1‰) свидетельствует о том. что они откладывались в приречной зоне поймы в условиях прирусловой отмели. Спорово-пыльцевые спектры характеризуются содержанием пыльцы древесных пород до 70% с преобладанием пыльцы березы (38–42%) и высоким содержанием пыльцы ели (до 18%) [10]. Пачка аллювия в интервале 1,96–0,35 м. представленная светло-серым мелкозернистым песком с прослоями аллохтонного торфа. характеризуется высокой льдистостью (до 50%). Спорово-пыльцевые спектры – типично тундрового типа. характеризуются преобладанием пыльцы кустарничков и трав (до 58%). Литологические особенности горизонта и высокое содержание водорастворимых солей (до 0,22%). позволяют предположить. что седиментация отложений происходила в заливе. образовавшемся в результате подтопления.
Во льду наиболее широкой ледяной жилы в устье р.Тамбей отмечено очень высокое для тундровых палиноспектров содержание пыльцы древесных пород – от 48 до 72% (табл. 3, рис. 11, а), что очевидно отражает состав весеннего пыльцевого дождя времени формирования ледяных жил. Наблюдается и смена доминантов: при сравнительно низком содержании пыльцы древесных пород доминирует береза, при высоком– ель. Это скорее всего свидетельствует о непрерывном накоплении льда жилы в различных ландшафтно-фациальных условиях. Особенность палиноспектров заметное участие пыльцы ели, которая в больших количествах по воздуху не переносится, скорее можно предположить ее перенос водой. Пыльца ели хорошей сохранности не отличается по цвету, но ее изменения ее содержания коррелируют колебаниями содержания переотложенных дочетвертичных форм. Очевидно, что в ледяные жилы пыльца ели и дочетвертичные пыльца и спор попадали из одного источника. Основным источником переотложенных палиноморф служат на Ямале служат отложения морского и лагунно-морского генезиса [11]. Поскольку в интервале 0.15–1.6 м выделена пачка песков лагунно-морского генезиса, вероятнее всего основное тело ледяной жилы накопилось при осаждении лагунно-морских отложений при уровне Обской губы выше современного. И, таким образом, особенности палиноспектров повторно-жильных льдов в долине р. Тамбей объяснимы их формированием в условиях периодического подтопления водами Обской губы. Сравнение с субфоссильными палиноспектрами показало, что в период формирования повторно-жильных льдов растительность Ямала существенно отличалась от современной. Условия для роста деревьев были в тот период даже в пределах современных арктических тундр.
Сходная картина наблюдается в палиноспектрах повторно-жильных льдов толщи первой террасы о. Белый. Здесь также наблюдается абсолютное доминирование пыльцы древесных пород (Picea 3-12%, Pinus sibirica 3-22%, Betula sect. Albae 23-40%). Участие морских вод в формировании льда непосредственно подтверждено результатами определения химического состава льда. Соленость изученных повторно-жильных льдов в 5-8 раз выше, чем в современных сингенетических жилах, формирующихся там же на низкой лайде. Представленные материалы показывают, что. спорово-пыльцевые спектры можно использовать для оценки фациальной составляющей. Анализ состава палиноспектров повторно-жильных льдов на о.Белом (табл. 4, рис. 11, б) показал, что у палиноспектров два источника – весенний пыльцевой дождь, попадавший на снег и отложения, вмещающие жилы. Второй источник менее существенен, поскольку в повторно-жильном льду практически не отмечено переотложенных древних пыльцы и спор, которые характерны для вмещающих отложений изученных льдов. Палиноспектры однородны – вероятно, накопление изученного льда происходило в стабильных климатических условиях, и возможно, повторно-жильные льды сформировались достаточно быстро. То, что в повторно-жильные льды попадал весенний пыльцевой дождь, подтверждается тем, что в них преобладает легко переносимая ветром пыльца хвойных пород и берез, обычно преобладающая в данном районе весной [12] а также пыльца других анемофильных растений – полыни. маревых. злаков. Низкое содержание пыльцы карликовой березки и ольховника говорит о том, что, когда в более южных районах начиналось их пыление, талая вода уже не проникала в трещины, т.к. к этому времени они закрывались. По той же причине практически отсутствует и пыльца травянистых растений.
Таблица 3. Состав пыльцы и спор в повторно-жильных льдах в пойменных отложениях в долине
р.Тамбей (%, в последней строке – экз/л)
|
номер образца
|
198-
YuV/1
|
198-
YuV/2
|
198-
YuV/3
|
198-
YuV/4
|
198-
YuV/6
|
198-
YuV/7
|
198-
YuV/20
|
|
Глубина, м
|
0.4
|
0.4
|
0.4
|
0.4
|
0.7
|
0.7
|
0.8
|
|
Пыльца деревьев
|
52
|
57
|
58
|
56
|
53
|
68
|
86
|
|
Пыльца кустарников
|
4
|
4
|
11
|
9
|
9
|
3
|
6
|
|
Пыльца трав и кустарничков
|
28
|
21
|
20
|
28
|
33
|
23
|
6
|
|
Споры
|
16
|
18
|
11
|
7
|
5
|
6
|
2
|
|
Larix
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Pinus sibirica
|
20
|
12
|
19
|
36
|
28
|
45
|
22
|
|
Pinus sylvestris
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Picea
|
20
|
8
|
22
|
20
|
9
|
10
|
58
|
|
Alnus
|
-
|
-
|
-
|
-
|
3
|
3
|
3
|
|
Betula
|
12
|
37
|
17
|
-
|
13
|
10
|
3
|
|
Betula sect. Nanae
|
4
|
4
|
9
|
9
|
9
|
3
|
3
|
|
Alnaster
|
-
|
-
|
2
|
-
|
-
|
-
|
3
|
|
Poaceae
|
4
|
2
|
14
|
-
|
4
|
10
|
3
|
|
Cyperaceae
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Ericaceae
|
4
|
6
|
2
|
13
|
23
|
-
|
-
|
|
Chenopodiaceae
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Artemisia
|
|
|
|
|
|
|
|
|
Varia
|
20
|
1
|
2
|
8
|
2
|
6
|
3
|
|
Bryales
|
8
|
6
|
3
|
-
|
-
|
3
|
2
|
|
Sphagnum sp.
|
8
|
7
|
8
|
2
|
5
|
3
|
-
|
|
Polypodiaceae
|
-
|
5
|
-
|
5
|
-
|
-
|
-
|
|
Lycopodium sp
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Экз/л
|
100
|
138
|
128
|
166
|
150
|
124
|
126
|
|
Переотложенные
|
11
|
4
|
12
|
8
|
12
|
24
|
8
|
|
номер образца
|
198-
YuV/21
|
198-
YuV/27
|
198-
YuV/8
|
198-
YuV/9
|
198-
YuV/10
|
198-
YuV/29
|
198-
YuV/15
|
|
Глубина.м
|
0.8
|
0.8
|
1.0
|
1.0
|
1.1
|
1.2
|
2.4
|
|
Пыльца деревьев
|
54
|
61
|
56
|
64
|
59
|
69
|
61
|
|
Пыльца кустарников
|
5
|
3
|
4
|
4
|
3
|
4
|
5
|
|
Пыльца трав и кустарничков
|
27
|
30
|
28
|
16
|
22
|
15
|
21
|
|
Споры
|
14
|
6
|
12
|
16
|
16
|
12
|
13
|
|
Larix
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Pinus sibirica
|
26
|
28
|
48
|
34
|
36
|
45
|
38
|
|
Pinus sylvestris
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Picea
|
7
|
6
|
4
|
18
|
17
|
11
|
13
|
|
Alnus
|
4
|
5
|
-
|
8
|
-
|
2
|
-
|
|
Betula
|
17
|
24
|
4
|
4
|
6
|
11
|
10
|
|
Betula sect. Nanae
|
5
|
-
|
4
|
4
|
-
|
4
|
5
|
|
Alnaster
|
-
|
3
|
-
|
-
|
3
|
-
|
-
|
|
Poaceae
|
13
|
16
|
4
|
8
|
10
|
11
|
7
|
|
Cyperaceae
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Ericaceae
|
2
|
3
|
-
|
-
|
6
|
-
|
3
|
|
Chenopodiaceae
|
-
|
-
|
-
|
-
|
--
|
-
|
-
|
|
Artemisia
|
3
|
8
|
20
|
4
|
3
|
2
|
3
|
|
Varia
|
9
|
3
|
4
|
4
|
3
|
2
|
8
|
|
Bryales
|
8
|
-
|
4
|
-
|
3
|
2
|
3
|
|
Sphagnum sp.
|
4
|
6
|
4
|
8
|
7
|
4
|
5
|
|
Polypodiaceae
|
2
|
-
|
4
|
8
|
6
|
6
|
5
|
|
Lycopodium sp
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Экз/л
|
117
|
72
|
100
|
103
|
123
|
116
|
68
|
|
Переотложенные
|
6
|
4
|
2
|
18
|
16
|
9
|
7
|
Таблица 4.
Содержание пыльцы и спор в повторно-жильном льду первой террасы острова Белый (%, в последней строке – экз/л)
|
Таксон
|
№ образца (глубина отбора. м)
|
|
YuV-209/1
(0.5)
|
YuV-209/2
(0.5)
|
YuV-209/3
(1.1)
|
YuV-209/4
(1.1)
|
YuV-209/5
(1.4)
|
|
Деревья
(пыльца). в т.ч.:
|
66.7
|
57.9
|
70.4
|
71.4
|
77.8
|
|
Picea sp.
|
10.0
|
15.8
|
3.7
|
14.3
|
2.8
|
|
Pinus sibirica
|
13.3
|
15.8
|
29.6
|
7.1
|
27.8
|
|
Pinus sylvestris
|
-
|
-
|
-
|
7.1
|
2.8
|
|
Alnus sp.
|
3.3
|
-
|
-
|
7.1
|
|
|
Betula sect. Albae
|
40.0
|
26.3
|
37.0
|
35.7
|
44.4
|
|
Кустарники
(пыльца). в т.ч.:
|
3.3
|
0
|
0
|
0
|
5.6
|
|
Salix sp.
|
-
|
-
|
-
|
-
|
2.8
|
|
Betula sect. Nanae
|
3.3
|
-
|
-
|
-
|
2.8
|
|
Травы и кустарнички
(пыльца). в т.ч.:
|
30.0
|
42.1
|
29.6
|
28.6
|
16.7
|
|
Poaceae
|
-
|
5.3
|
3.7
|
21.4
|
5.6
|
|
Cyperaceae
|
-
|
-
|
-
|
7.1
|
2.8
|
|
Ericaceae
|
-
|
-
|
3.7
|
-
|
-
|
|
Chenopodiaceae
|
6.7
|
10.5
|
14.8
|
-
|
2.8
|
|
Artemisia sp.
|
13.3
|
21.1
|
7.4
|
-
|
5.6
|
|
Ranunculaceae
|
6.7
|
-
|
-
|
-
|
-
|
|
Fabaceae
|
3.3
|
5.3
|
-
|
-
|
-
|
|
Мхи и папоротники
(споры). в т.ч.:
|
23.1
|
24.0
|
18.2
|
39.1
|
14.3
|
|
Bryales
|
12.8
|
6.8
|
6.1
|
21.7
|
11.9
|
|
Sphagnum sp.
|
7.7
|
6.8
|
6.1
|
13.0
|
-
|
|
Polypodiaceae
|
2.6
|
-
|
6.1
|
4.3
|
2.4
|
|
Концентрация. экз./л*
|
207
|
132
|
180
|
111
|
234
|
*.
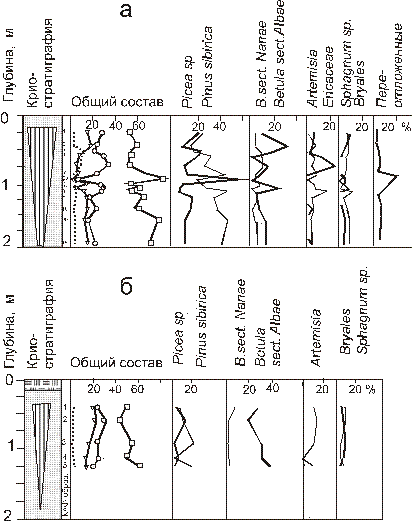
Рис. 11. Спорово-пыльцевая диаграмма из повторно-жильных льдов. в толще пойменных отложений р. Тамбей на северо-востоке полуострова Ямал (а) и отложений первой морской террасы о. Белый (б). 1 – торф. 2 – песок. 3 – повторно-жильный лед. 4 – пыльца древесных пород. 5 – пыльца кустарников. 6 – пыльца трав и кустарничков. 7 – споры; номера образцов соответствуют таблицам 4,5
ДИСКУССИЯ
Согласно макроциклической модели формирования мощных сингенетических повторно-жильных льдов Ю.К. Васильчука [13], основной рост ледяных жил происходит во время субаэральной фазы, когда накапливается торф или оторфованные отложения, в этот момент в повторно-жильный лед попадает в основном пыльца, принесенная ветром на поверхность снежного покрова ранней весной. Периодически режим сменяется субаквальным, когда накапливаются гравий, песок, супесь, суглинки, глины. В это время в жилы может проникать пыльца и споры, содержащиеся в морских и речных водах. Смена субаэрального режима субаквальным может быть связана с подпруживанием небольших рек, подтоплением пойм, опусканием побережий морей, заливов и губ, образованием запруд или обширных соров, часто с размывом ранее накопившихся отложений. В этот период жилы консервируются или накапливаются очень слабо, в палиноспектрах, относящихся к этому моменту, могут появиться дочетвертичные пыльца и споры, а также очень высокое, не соответствующее региональным фоновым значениям содержание пыльцы древесных пород.
Сравнивая палиноспектры повторно-жильных льдов на острове Белом и в долине реки Тамбей отметим их сходство и отличие от современных палиноспектров из этих же районов. Это, в первую очередь, высокое содержание пыльцы древесных пород 57,9-77,8% на о Белом и 52-86% в долине р.Тамбей (табл. 5). Примерно одинаковый состав таксонов, за исключением того, что в повторно-жильных льдах в долине р.Тамбей не отмечена пыльца сосны обыкновенной и осок, в то время как на о.Белом содержание пыльцы этих таксонов колеблется от 0 до 7%. Основное отличие в содержании пыльцы ели, которое во льду в долине р.Тамбей достигает 58%. Очевидно, что повторно-жильные льды накапливались в период благоприятный для существования древесной растительности севернее ее современного распространения, но под воздействием разных факторов. Если на острове Белом пыльца и споры в основном перенесены ветром, при этом отмечаются компоненты, такие как пыльца лютиковых, бобовых, осок, которая как правило выпадает на поверхность в середине лета. Наличие более минерализованных клиньев и летних компонентов приводит к предположению, что повторно-жильные льды формировались в условиях мелких озер, в которые летом попадала пыльца трав, а основной палинологический сигнал формировался из весеннего пыльцевого дождя, т.е. в трещины попадала пыльца, выпавшая на снег до того, как трещина заполнилась. В долине р.Тамбей помимо весеннего пыльцевого дождя поступлению пыльцы и спор в повторно-жильный лед способствовало подтопление водами Обской губы. Содержание пыльцы кустарников ольховника и карликовой березки, вероятно, отражает участие в растительном покрове. Следовательно, если в формировании повторно-жильных льдов в долине р.Тамбей в ее приустьевой части отмечено участие вод Обской губы, то на острове Белом повторно-жильные льды вероятнее всего формировались в условиях мелких водоемов с повышенным содержанием морских солей. Этот процесс и сейчас типичен на о. Белом (рис. 12).
Таблица 5.
Диапазон значений содержания пыльцы и спор (%) и концентрации (экз./л) в голоценовых повторно-жильных льдах в долине р.Тамбей и на о. Белом
|
Таксон
|
Первая терраса о.Белый
|
Высокая пойма р.Тамбей
|
|
пыльца деревьев
|
57.9-77.8
|
52-86
|
|
Picea sp.
|
2.8-15.8
|
6-58
|
|
Pinus sibirica
|
7.1-29.6
|
12-45
|
|
Pinus sylvestris
|
0-7.1
|
-
|
|
Alnus sp.
|
0-7.1
|
0-3
|
|
Betula sect. Albae
|
26.3-44.4
|
0-48
|
|
Пыльца кустарников
|
0-5.6
|
3-11
|
|
Salix sp.
|
0-2.8-
|
-
|
|
Betula sect. Nanae
|
0-3.3
|
0-9
|
|
Пыльца трав и кустарничков
|
16.7-42.1
|
6-33
|
|
Poaceae
|
0-21.4-
|
0-16
|
|
Cyperaceae
|
0-7.1
|
-
|
|
Ericaceae
|
0-3.7
|
0-23
|
|
Chenopodiaceae
|
0-14.8
|
-
|
|
Artemisia sp.
|
0-21.1
|
0-20
|
|
Ranunculaceae
|
0-6.7
|
0-2-
|
|
Fabaceae
|
0-5.3
|
0-4
|
|
споры:
|
14.3-39.1
|
2-18
|
|
Bryales
|
6.1-21.7
|
0-8
|
|
Sphagnum sp.
|
0-13.0
|
0-8
|
|
Polypodiaceae
|
0-6.1
|
0-8
|
|
Концентрация. экз./л
|
111-234
|
68-123
|

Рис. 12. Развитие повторно-жильных льдов на о. Белом
Полигоны прослеживаются как на поверхности морских террас (а), так и на дне мелких озер (б) (фото П.Т. Орехова) [14]
Решение вопроса о периоде формирования повторно-жильных льдов в эпоху существенного улучшения условий вегетации определено полученными в разное время радиоуглеродными датировками древесины из этих районов, палинологическими данными, а также результатами исследований более северных территорий. Характерным процессом для бореального периода были сравнительно интенсивные перемещения береговой линии, относительно быстрое повышение уровня моря до отметок + 8-10 м по сравнению с современными, с чем связано накопление основной толщи отложений первых террас в низовьях рек и вдоль побережий [15]. При переходе к «оптимальной» фазе бореального периода началось снижение уровня моря, что привело к возникновению на поверхности первых террас остаточных водоемов. По нашему мнению, именно в таких засоленных мелководных водоемах сформировались повторно-жильные льды на о.Белом. Состав палиноспектров определялся принципиально другим распределением древесной растительности. По нашим данным на основании около 200 радиоуглеродных датировок древесины отобранной в пределах п-ова Ямал [15, 16] распространение древесной растительности в пределах территории современной тундры датируется от 9100–9200 лет назад до примерно 4900 лет назад (рис. 13). Не удивительно, что палиноспектры в долине р.Тамбей и на о. Белом содержат значительное количество пыльцы древесных пород
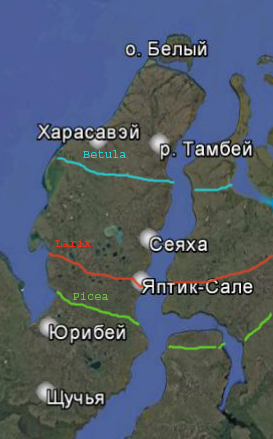
Рис. 13. Северные границы ареалов ели, березы, и лиственницы по данным Vasil'chuk et al. [16] и MacDonald et al. [17]
В верхней части Сеяхинской едомы была получена датировка 11620 ± 90 лет (Hel-3942). Палиноспектры, из отложений над едомой отражают повышение концентрации пыльцы хвойных сосны обыкновенной и ели. Датировки древесины на севере полуострова Ямал получены на р. Пухучаяха 8250 ±80 (ЛУ-1139); 9300 ±100 (ГИН-2442); устье р. Сёяха-Зеленая (Вост. Ямал), 9190±80 (ЛУ-1086); р. Нябыяха (вост. Ямал), 9160±120(ЛУ-714); оз. Малто (Центр. Ямал). В северной части Енисейского залива [18] в отложениях датированных 8285 ± 40 -7870 ± 40 выделяется смена максимума содержания пыльцы березы максимумом ели. На это время приходится самая высокая концентрация пыльцы и спор. Для севера Западной Сибири – это наиболее благоприятные условия вегетационного периода. Например, в период около 8300-8200 лет на границе гипоарктических и арктических тундр возникла березовая роща в районе устья р. Сеяха (Зеленая). Возможно, возникновение растительных формаций этого типа связано с условиями, сходными с климатом позднего плейстоцена, когда отсутствие облачности приводило к повышению летних температур и к росту изолированных рощ в наиболее благоприятных условиях, по тип современных лесотундр.
В зоне полярных пустынь также прослеживается ранне голоценовое повышение летних температур наряду с формированием повторно-жильных. Палинологические данные получены П.Е. Тарасовым и А.А. Андреевым по погребенному торфянику, изученному Ф.А. Романенко на о. Свердруп [19]. Горизонт погребенного торфа с остатками карликовой березки залегает в интервале 0,3-0,9 м. его основание на глубине 0,8 - 0,9 м. датировано 11640 ± 40 лет назад (ГИН-7625). Судя по составу локальных компонентов спорово-пыльцевых спектров растительность того времени на о.Свердруп отвечала палиноспектрам южной полосы арктических тундр с участием Salix, Ranunculaceae, Rosaceae, Polygonum, Caryophyllaceae, Valeriana, Lycopodium alpinum, L.selago, Bryales. Региональный пыльцевой дождь был представлен Poaceae, Cyperaceae, Artemisia, Asteraceae, Betula sect. Nanae, Salix, Alnaster. Отмечена также дальнезаносная пыльца Pinus sylvestris, P.sibirica, Picea obovata, Betula sect. Albae. Содержание пыльцы этих древесных пород в несколько раз меньше, чем в субфоссильном спектре на о.Свердруп.
Выводы:
Результаты рассмотрения данных по повторно-жильным льдам в долине р.Тамбей и на о. Белом подтверждают существенное распространение древесной растительности на Ямале в раннем голоцене. Поскольку в повторно-жильных льдах зафиксирован весенний пыльцевой дождь, эти данные отражают более северное положение границы распространения древесных пород. Повышение летних температур в раннем голоцене не приводило к снижению интенсивности повторно-жильного льдообразования, т.е - это период усиления континентальности климата.
На составе палиноспектров повторно-жильных льдов о. Белый и долины р.Тамбей отразилось воздействие разных факторов: в первом случае формирование повторно-жильных льдов в условиях мелких озер, что давало возможность попадания в лед воды засоленного водоемы и пыльцы и спор, попавших в этот водоем в летний сезон; во втором случае, дополнительным фактором послужила неоднократная кратковременная ингрессия вод Обской губы, это определило повышенное содержание пыльцы ели в палиноспектрах повторно-жильных льдов.
Библиография
1. Ильина И.С., Лапшина Е.И., Лавренко Н.Н., Мельцер Л.И., Романова Е.А., Богоявленский Б.А., Махно В.Д. Растительный покров Западно-Сибирской равнины / ред. Воробьев В.В., Белов А.В. Новосибирск: Наука. Сиб. отд. 1985. 251 c.
2. Трофимов В.Т., Баду Ю.Б., Кудряшов В.Г., Фирсов Н.Г. Полуостров Ямал. М.: Изд-во МГУ. 1975. 278 с.
3. Природа Ямала. Екатеринбург: Наука. 1995. 436 с.
4. Андреев М.П. Лишайники арктических тундр верховьев р. Тамбей (полуостров Ямал) // Новости систематики низших растений. 1982. Т. 19. С. 111-116.
5. Жадринская Н.Г. Растительный покров // Ямало-Гыданская область (физико-географическая характеристика). Л.. 1977. С. 168-172.
6. Тишков А. А., Кренке А. Н. «Позеленение» Арктики в ХХI в. как эффект синергизма действия глобального потепления и хозяйственного освоения // Арктика: экология и экономика № 4 (20). 2015. С.28-37
7. Васильчук А.К. Региональная и дальнезаносная пыльца в тундровых палиноспектрах // Известия Российской Академии Наук. Серия биологическая. 2005. №1. С. 85–99.
8. Васильчук А.К., Васильчук Ю.К. Инженерно-геологические и геохимические условия полигональных ландшафтов в районе устья реки Тамбей (север полуострова Ямал)// Инженерная геология. 2015. № 4. С. 36-54.
9. Савельева Л.А., Рашке (Морозова) Е.А.,Титова Д.В. Атлас фотографий растений и пыльцы дельты реки Лены. СПб.: тип.«Ренова», 2013. 114 с.
10. Васильчук А.К., Васильчук Ю.К. Инженерно-геологические и геохимические условия полигональных ландшафтов в районе устья реки Тамбей (север полуострова Ямал) // Инженерная геология. 2015. № 4. С. 36-54
11. Васильчук А.К. Особенности тафономии пыльцы и спор в Арктике // Известия Российской Академии Наук. Серия биологическая. 2005. №2. С. 240–252.
12. Васильчук А.К. Региональная и дальнезаносная пыльца в тундровых палиноспектрах // Известия Российской Академии Наук. Серия биологическая. 2005. №1. С. 85 – 99.
13. Васильчук Ю.К. Повторно-жильные льды; гетероцикличность. гетерохронность. гетерогенность М.: Изд-во Московского университета. 2006. 404 c.
14. Walker D.A., Orekhov P., Frost G.V., Matyshak G., Epstein H.E., Leibman M.O., Khitun O., Khomotov A., Daanen R., Gobroski K., Maier H.A. The 2009 Yamal expedition to ostrov Belyy and Kharp, Yamal region, Russia. Data report. Alaska Geobotany Center, Institute of Arctic Biology, University of Alaska Fairbanks, AK. 2009. 49 p.
15. Васильчук Ю.К., Петрова Е.А., Васильчук А.К. Некоторые черты палеогеографии голоцена Ямала // Бюллетень Комиссии по изучению четвертичного периода. 1983. Вып. 52. С. 73–89
16. Vasil'chuk Yu.K., Jungner H., Vasil'chuk A.C. 14C dating of peat and δ18O-δD in ground ice from Northwest Siberia // Radiocarbon. 2001. Vol.43. N 2B. P. 527–540.
17. MacDonald G.M., Velichko A.A., Kremenetski C.V., Borisova O.K., Goleva A.A., Andreev A.A., Cwynar L.C., Riding R.T., Forman S.L., Edwards T.W.D., Aravena R., Hammarlund D., Szeicz Ju.M., Gattaulin V.N. Holocene Treeline History and Climate Change Across Northern Eurasia // Quaternary Research. 2000. Vol.53. P. 302–311.
18. Kraus M., Matthiessen J., Stein R. A Holocene marine pollen record from the northern Yenisei Estuary (southeastern Kara Sea, Siberia) // Proceedings in Marine Science. Siberian River run-off in the Kara Sea: characterization, quantification, variability and environmental significance. Eds.: R.Stein, K.Fahl., D.K.Futterer, E.Galimov. 2003. Vol. 6. P. 435–456.
19. Тарасов П.Е., Андреев А.А., Романенко Ф.А., Сулержицкий Л.Д. Палиностратиграфия верхнечетвертичных отложений острова Свердруп (Карское море) // Стратиграфия. Геологическая корреляция. 1995. Том 3. №2. С. 98–104.
References
1. Il'ina I.S., Lapshina E.I., Lavrenko N.N., Mel'tser L.I., Romanova E.A., Bogoyavlenskii B.A., Makhno V.D. Rastitel'nyi pokrov Zapadno-Sibirskoi ravniny / red. Vorob'ev V.V., Belov A.V. Novosibirsk: Nauka. Sib. otd. 1985. 251 c.
2. Trofimov V.T., Badu Yu.B., Kudryashov V.G., Firsov N.G. Poluostrov Yamal. M.: Izd-vo MGU. 1975. 278 s.
3. Priroda Yamala. Ekaterinburg: Nauka. 1995. 436 s.
4. Andreev M.P. Lishainiki arkticheskikh tundr verkhov'ev r. Tambei (poluostrov Yamal) // Novosti sistematiki nizshikh rastenii. 1982. T. 19. S. 111-116.
5. Zhadrinskaya N.G. Rastitel'nyi pokrov // Yamalo-Gydanskaya oblast' (fiziko-geograficheskaya kharakteristika). L.. 1977. S. 168-172.
6. Tishkov A. A., Krenke A. N. «Pozelenenie» Arktiki v KhKhI v. kak effekt sinergizma deistviya global'nogo potepleniya i khozyaistvennogo osvoeniya // Arktika: ekologiya i ekonomika № 4 (20). 2015. S.28-37
7. Vasil'chuk A.K. Regional'naya i dal'nezanosnaya pyl'tsa v tundrovykh palinospektrakh // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2005. №1. S. 85–99.
8. Vasil'chuk A.K., Vasil'chuk Yu.K. Inzhenerno-geologicheskie i geokhimicheskie usloviya poligonal'nykh landshaftov v raione ust'ya reki Tambei (sever poluostrova Yamal)// Inzhenernaya geologiya. 2015. № 4. S. 36-54.
9. Savel'eva L.A., Rashke (Morozova) E.A.,Titova D.V. Atlas fotografii rastenii i pyl'tsy del'ty reki Leny. SPb.: tip.«Renova», 2013. 114 s.
10. Vasil'chuk A.K., Vasil'chuk Yu.K. Inzhenerno-geologicheskie i geokhimicheskie usloviya poligonal'nykh landshaftov v raione ust'ya reki Tambei (sever poluostrova Yamal) // Inzhenernaya geologiya. 2015. № 4. S. 36-54
11. Vasil'chuk A.K. Osobennosti tafonomii pyl'tsy i spor v Arktike // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2005. №2. S. 240–252.
12. Vasil'chuk A.K. Regional'naya i dal'nezanosnaya pyl'tsa v tundrovykh palinospektrakh // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. 2005. №1. S. 85 – 99.
13. Vasil'chuk Yu.K. Povtorno-zhil'nye l'dy; geterotsiklichnost'. geterokhronnost'. geterogennost' M.: Izd-vo Moskovskogo universiteta. 2006. 404 c.
14. Walker D.A., Orekhov P., Frost G.V., Matyshak G., Epstein H.E., Leibman M.O., Khitun O., Khomotov A., Daanen R., Gobroski K., Maier H.A. The 2009 Yamal expedition to ostrov Belyy and Kharp, Yamal region, Russia. Data report. Alaska Geobotany Center, Institute of Arctic Biology, University of Alaska Fairbanks, AK. 2009. 49 p.
15. Vasil'chuk Yu.K., Petrova E.A., Vasil'chuk A.K. Nekotorye cherty paleogeografii golotsena Yamala // Byulleten' Komissii po izucheniyu chetvertichnogo perioda. 1983. Vyp. 52. S. 73–89
16. Vasil'chuk Yu.K., Jungner H., Vasil'chuk A.C. 14C dating of peat and δ18O-δD in ground ice from Northwest Siberia // Radiocarbon. 2001. Vol.43. N 2B. P. 527–540.
17. MacDonald G.M., Velichko A.A., Kremenetski C.V., Borisova O.K., Goleva A.A., Andreev A.A., Cwynar L.C., Riding R.T., Forman S.L., Edwards T.W.D., Aravena R., Hammarlund D., Szeicz Ju.M., Gattaulin V.N. Holocene Treeline History and Climate Change Across Northern Eurasia // Quaternary Research. 2000. Vol.53. P. 302–311.
18. Kraus M., Matthiessen J., Stein R. A Holocene marine pollen record from the northern Yenisei Estuary (southeastern Kara Sea, Siberia) // Proceedings in Marine Science. Siberian River run-off in the Kara Sea: characterization, quantification, variability and environmental significance. Eds.: R.Stein, K.Fahl., D.K.Futterer, E.Galimov. 2003. Vol. 6. P. 435–456.
19. Tarasov P.E., Andreev A.A., Romanenko F.A., Sulerzhitskii L.D. Palinostratigrafiya verkhnechetvertichnykh otlozhenii ostrova Sverdrup (Karskoe more) // Stratigrafiya. Geologicheskaya korrelyatsiya. 1995. Tom 3. №2. S. 98–104.
|















